
Nierstraszella andamanica ( Smith, 1906 ) Sigwart, 2009
|
publication ID |
https://doi.org/ 10.1080/00222930802604157 |
|
persistent identifier |
https://treatment.plazi.org/id/5F5C879D-FFCC-F40F-4722-8248F3B3FB09 |
|
treatment provided by |
Felipe |
|
scientific name |
Nierstraszella andamanica ( Smith, 1906 ) |
| status |
comb. nov. |
Nierstraszella andamanica ( Smith, 1906) View in CoL comb. nov., nom. rev.
Chresonymy and synonymy revised from Saito (1997), Sirenko (1992), and Schwabe et al. (2008).
Lepidopleurus andamanicus Smith, 1906, p. 251 [lectotype BMNH 1906.10.12.86 (designated by Kaas and Van Belle 1985: 118), three paralectotypes BMNH 1906.10.12.87–89, paralectotype pro parte (girdle preparation) RMNH MOL.K.5024; type locality: India, Andaman Island off North Sentinel Island (11 ° 339 N 92 ° 159 E), 240 fathoms (439 m)]; Annandale and Stewart 1909, figures 4, 4a; Winckworth, 1940, p. 19; Rajagopal and Subba Rao 1974, p. 400; Schwabe 2006b, p. 20.
Lepidopleurus porosus Leloup, 1981, p. 322 , figures 5–6, pl. 2, figures 4–6 (holotype MNHN 6012, type locality: Philippines, 13 ° 40.79 N 120 ° 309 E, Campagne MUSORSTOM 1: stn CP47, 685– 757 m); Kaas and Van Belle 1985, p. 126; Kaas and VanBelle 1998, p. 148.
Lepidopleurus philippinus Leloup, 1981, p. 322 , figure 4, pl. 2, figures 1–3 (holotype MNHN 5981, paratypes MNHN 5979, 5980, 5988, type locality: Philippines, 13 ° 50.59 N 120 ° 289 E, Campagne MUSORSTOM 1: stn CP43, 448– 484 m); Sirenko 1992, p. 84.
Leptochiton andamanicus: Kaas 1982, p. 89 ; Kaas and Van Belle 1980, p. 7; Kaas and Van Belle 1998, p. 19.
Leptochiton philippinus: Kaas and Van Belle 1998, p. 143 .
Nierstraszella lineata View in CoL e.p.: Sirenko 1992, p. 84.
Nierstraszella philippina: d’Hondt 2006, p. 258 View in CoL .
Type material
BMNH 1906.10 .12.86 (lectotype, designated by Kaas and Van Belle 1985); 1906.10.12.87-89 (three paralectotypes) ; RMNH MOL.K.5024 (paralectotype pro parte, girdle preparation).
Type locality
Bay of Bengal , off North Sentinel Island, Andaman Islands group, 432 m.
Material examined
Lepidopleurus andamanicus lectotype BMNH 1906.10.12.86; L. andamanicus paralectotypes BMNH 1906.10.12.87-89; L. andamanicus paralectotype pro parte RMNH MOL.K.5024; Lepidopleurus philippinus holotype (MNHN 5981) and paratypes (MNHN); Lepidopleurus niasicus holotype ( ZMB); Lepidopleurus porosus holotype (MNHN 6012); RMNH MOL.HLS.1748; RMNH MOL.K.4966; for new expedition material from the West Pacific please see Appendix 1 .
Diagnosis
As for genus. Periostracum without pustules, sometimes abraded on older area of valve. Aesthete caps visible or abraded, forming impression of regularly spaced pores. Each megalaesthete with two rows of micraesthetes, six or seven each side. Girdle covered in small blunt-pointed spicules. Gills increasing in number with the size of the animal, eight per side in juvenile specimens (1.7 mm wide) to 15–20 per side in adults.
Description
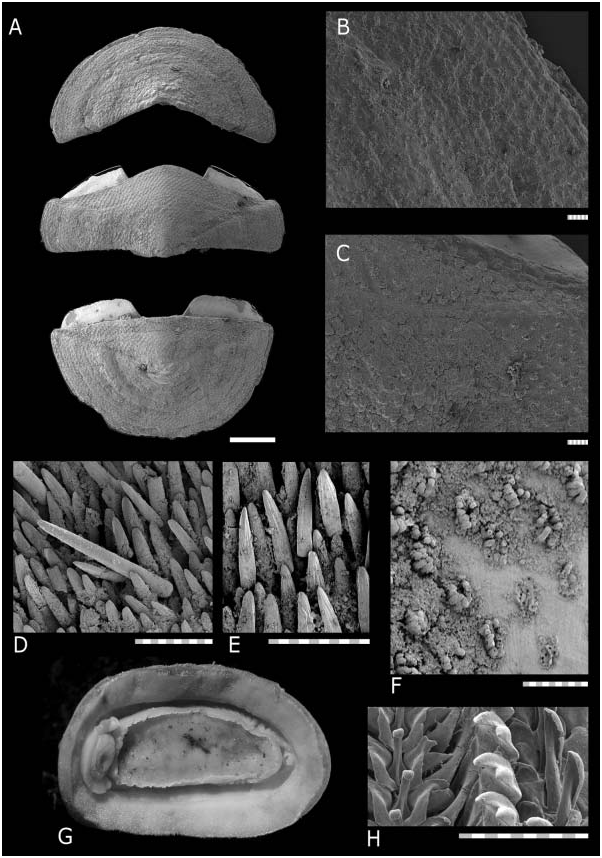
Animal up to 27 × 13 mm long (‘‘Salomon 2’’ CP 2228). Valves carinated, moderately elevated (dorsal elevation ratio approx. 0.5 on valve II), valves distinctly beaked.
armature, specimen from Solomon Islands, ‘‘Salomon 2’’ sta. CP2280; (E), ventral girdle armature, specimen from ‘‘Salomon 2’’ sta. CP2264; (F) partially abraded section of intermediate valve dorsal surface showing aesthete caps on individual aesthete clusters, specimen from ‘‘Salomon 2’’ sta. CP2264; (G) ventral view of preserved animal, anterior at left, specimen from ‘‘Salomon 2’’ sta. CP2264; (H) radula, showing half of central area of radular row, specimen RMNH MOL.K.4966. (A–F) Anterior is at top; (B–F,H) scale bars 100 mm.
Head valve semicircular, slightly narrower than tail valve. Intermediate valves rectangular, lateral areas distinct but little inflated, and not depressed near apices. Anterior margin slightly convex, and posterior margin concave around projecting apex, side margins straight.
Tegmentum smooth, without sculpture, covered in thick periostracum. Aesthete caps clearly visible, protruding from aesthete openings. Aesthete pores arranged quincuncially, sometimes giving an appearance of sculpture. Periostracum sometimes abraded; where thin, aesthete openings are visible as pores or slightly discoloured points. Colour of tegmentum white, older parts of valves (near apex) covered with black mineral deposits, sometimes extending to cover dorsal surface of entire animal.
Aesthete pores arranged in quincunx, with one megalaesthete with micraesthetes in rows on either side, approximately six per side. Megalaesthete 5 mm in diameter.
Articulamentum well developed; apophyses short and broad, widely separated, bluntly triangular in valves II–VII, round to trapezoidal in tail valve. Articulamentum forming flat thickened ridge along outer margins of end valves.
Girdle narrow, dorsally densely covered in elongate, bluntly pointed spicules (120 × 16 mm), with approximately five ribs covering entire length of spicule. Intersegmental areas with larger scales. Marginal fringe absent. Ventrally, girdle covered with elongate, flat, narrow scales with approximately five ribs as in dorsal scales (64 × 16 mm).
Radula major lateral teeth with bicuspid head; interior denticle shortest. In juvenile specimens, major lateral teeth are tricuspid (juvenile specimen examined 2.3 mm wide, approximately 4.1 mm long; smallest adult specimen with bicuspid major lateral cusps 6.5 mm wide, approximately 11.7 mm long).
In smallest juvenile specimens, there are eight gills per side (animal width 1.7 mm, eight gills, ‘‘Salomon 2’’ CP 2263). Number of gills increases with size of animal, up to 20 per side (animal width 10.4 mm, ‘‘Salomon 2’’ CP 2219), usually 16 gills per side in animals of width 5 mm or more.
Distribution
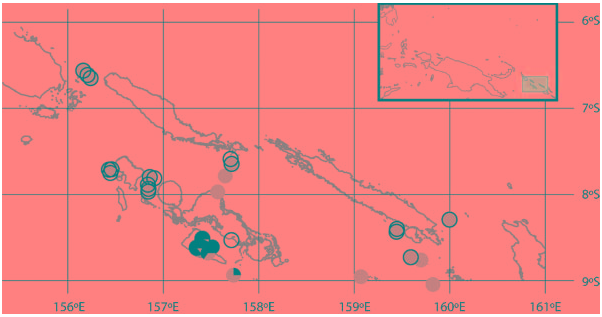
Widely distributed in the South Pacific, in the Philippines, Andaman Islands, Indonesia, Solomon Islands, and Vanuatu; from 177 to 1760 m.
Remarks
The name Lepidopleurus andamanicus Smith, 1906 has been selected as the earliest available name for this species. Lepidopleurus porosus Leloup, 1981 and Lepidopleurus philippinus Leloup, 1981 are junior synonyms, and have previously been recognized as having strong affinities with L. andamanicus . Kaas and Van Belle (1985, p. 128) noted that ‘‘ L. philippinus bears a close resemblance with L. andamanicus (Smith) . In fact the two are so similar that we first were apt to believe them to be conspecific.’’ In their redescription of Lepidopleurus philippinus Kaas and Van Belle (1985, p. 128) also noted ‘‘The unique type of L. porosus …is in all respects identical with philippinus .’’
Examination of the morphology, and particularly of the aesthetes of the present material, shows that there are in fact two distinct species of Nierstraszella , which correspond to Lepidopleurus lineatus and L. andamanicus ( Figures 1 View Figure 1 , 4 View Figure 4 ). Before this study few specimens of L. andamanicus had been collected, whereas L. lineatus is locally abundant in many sites throughout the Indo-West Pacific (e.g. Saito 2001). Previous descriptions of ‘‘ Nierstraszella ’’ sp. therefore correspond to accurate reports of L. lineatus .
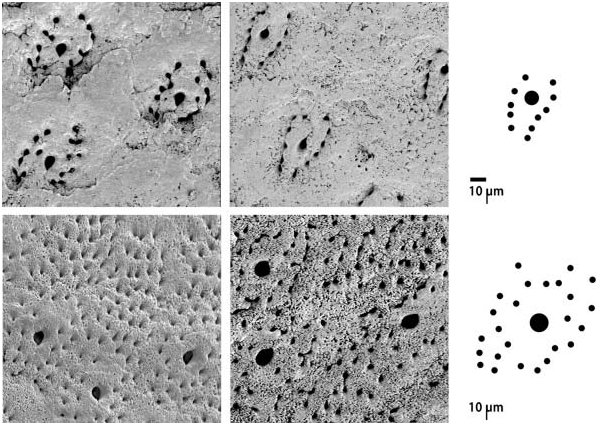
Lepidopleurus andamanicus does not have raised pustules and the aesthetes are arranged in discrete groups with a single megalaesthete, approximately 5 mm wide, surrounded by approximately 12 micraesthetes arranged in two rows, on either side of the central megalaesthete. Lepidopleurus lineatus , by contrast, has a distinctive pattern of randomly dispersed, very large megalaesthetes (approximately 10 mm wide), with scattered micraesthetes over the whole valve surface underneath large pustules ( Figures 2 View Figure 2 , 3F View Figure 3 ). During the course of this study it was not possible to examine name-bearing type material by SEM imaging, which would be required to see the pattern of aesthete arrangement in the two species. However, L. andamanicus is distinguished by several macroscopic features which clearly separate this species from L. lineatus and which have been compared on the type specimens examined.
Nierstraszella andamanica can be distinguished by its valves having higher elevation, more pronounced apex, and lacking the large raised pustules distinctive of L. lineatus . The periostracum in L. andamanicus usually does not form ‘‘sculpture’’ but in some cases where the periostracum is very thin small pustules are formed over aesthete caps, arranged in widespread quincunx ( Figure 4A View Figure 4 ); the periostracum and any appearance of sculpture always rub off easily with scraping ( Figure 4F View Figure 4 ). The shape of valve VIII is closer to semicircular in L. andamanicus , and the mucro is slightly anterior; in L. lineatus the tail valve appears larger and the mucro is medial. The girdle typically appears narrower in L. andamanicus and quite wide in L. lineatus .
Epibiotic bryozoans
The bryozoan Pseudobathyalozoon profundum colonizes the ventral girdle of the chitons, with the zooids invading the pallial cavity of the host ( Figure 5 View Figure 5 ). Pseudobathyalozoon preferentially colonizes N. andamanica and is less frequently found associated with the congener N. lineata . In material from the Philippines, N. andamanica was the only host organism colonized with the bryozoan. However, in the Solomon Islands, the bryozoan has been found on a small number of specimens of N. lineata (three specimens in two stations; ‘‘Salomon 2’’ CP2280, CP2273) and on one specimen of the chiton Ferreiraella plana ( Nierstrasz, 1905) (‘‘Salomon 2’’ CP2289). In two stations (CP2280, CP2289), the bryozoan Pseudobathyalozoon was found on these additional host species where N. andamanica was abundant and the majority of chiton specimens collected in a sample were colonized; in the third station (CP2273) the single specimen of N. lineata was one of three individual chitons collected.
In total, 307 specimens of N. andamanica were examined from the Philippines and Solomon Islands. Of those, 35% (n 5107) hosted epibiotic bryozoans. However, colonization was not present in all collecting stations, and was limited to localized areas in both the Philippines ( Figure 6 View Figure 6 ) and the Solomon Islands ( Figure 7 View Figure 7 ). In the Solomon Islands, areas where the epibiotic bryozoan is present show that 84% of specimens of N. andamanica are colonized; in the Philippines, however, only 59% of chitons in colonized areas carry the bryozoans ( Table 1).
In the Solomon Islands, animals ranged from 0.5 to 12.9 mm in width, with the smallest individual colonized at 2.3 mm wide. Within collecting stations where the colonizing bryozoan was present, the largest individual in the station was always colonized, with the rate of colonization increasing in larger size classes ( Table 2). For specimens of N. andamanica collected in these stations where Pseudobathyalozoon was present, there was a significant relationship between size of host and the presence of Pseudobathyalozoon zooids (x 2 531.9; df54; p,0.001). The largest individual chitons overall occurred in stations where there was no colonization, but the difference in size was not great (12.9 mm wide, compared with 11.9 mm for colonized chitons; both approximately 23 mm long).
The two populations of the epibiotic bryozoan have very different depth profiles and occur much deeper in the Philippines than in the Solomon Islands ( Table 1).
The bryozoans are attached all along the pallial cavity, on the ventral side of the girdle. Although quantitative data are not available for the distribution of the bryozoans on the host chitons, a number of points are readily apparent. Most
in (A); (D) dorsal girdle armature, specimen from Solomon Islands, ‘‘Salomon 2’’ sta. CP2243; (E) ventral girdle armature, specimen from ‘‘Salomon 2’’ sta. CP2243; (F) intermediate valve dorsal surface showing aesthete caps on individual aesthete clusters, specimen from ‘‘Salomon 2’’ sta. CP2226; (G) ventral view of preserved animal, anterior at left, specimen from ‘‘Salomon 2’’ sta. CP2243; (H) radula, showing half of central area of radular row, specimen RMNH MOL.HLS.2010. (A–F) Anterior is at top; (B–F,H) scale bars 100 mm.
bryozoan specimens were attached at the posterior half of the host, invading the space occupied by the host’s gills ( Figure 5 View Figure 5 ). A number were also found around the host’s head. Relatively few bryozoans attached along the intermediate part of the host body, along the anterior half of the foot, although bryozoans were occasionally present at all points on the pallial cavity. Bryozoans always attach basally to the ventral girdle and never to the pallial groove. There is no apparent preference for left or right side of the host overall, although bryozoans on an individual host may be either restricted to a small area or distributed along the entire length of the pallial groove.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nierstraszella andamanica ( Smith, 1906 )
| Sigwart, Julia D. 2009 |
Nierstraszella philippina: d’Hondt 2006 , p. 258
| d'Hondt J-L 2006: 258 |
Leptochiton philippinus:
| Kaas P & Van Belle RA 1998: 143 |
Nierstraszella lineata
| Sirenko BI 1992: 84 |
Leptochiton andamanicus:
| Kaas P & Van Belle RA 1998: 19 |
| Kaas P 1982: 89 |
| Kaas P & Van Belle RA 1980: 7 |
Lepidopleurus porosus
| Kaas P & Van Belle RA 1985: 126 |
| Leloup E 1981: 322 |
Lepidopleurus philippinus
| Sirenko BI 1992: 84 |
| Leloup E 1981: 322 |
Lepidopleurus andamanicus
| Schwabe E 2006: 20 |
| Kaas P & Van Belle RA 1985: 118 |
| Rajagopal S & Subba Rao NV 1974: 400 |
| Winckworth R 1940: 19 |
| Smith EA 1906: 251 |