
Palimphemus anceps Kner, 1862
|
publication ID |
https://doi.org/ 10.5252/g2012n3a9 |
|
persistent identifier |
https://treatment.plazi.org/id/626BBA02-FFA5-FFB7-27D5-AA8EFDDF358E |
|
treatment provided by |
Marcus |
|
scientific name |
Palimphemus anceps Kner, 1862 |
| status |
|
Palimphemus anceps Kner, 1862 ( Figs 1-8 View FIG View FIG View FIG View FIG View FIG ; 9A, D, F View FIG )
Palimphemus anceps Kner, 1862: 490-495 ; pl. 1, fig. 2; 1863: 148. — Woodward 1901: 270. — Schubert 1906: 697. — Schultz in Brzobohatý & Schultz 1978: 444. — Hofmann 2001: 221. — Schultz in Schmid, Harzhauser & Kroh 2001: 22.
HOLOTYPE. — PIWU, without catalogue number.
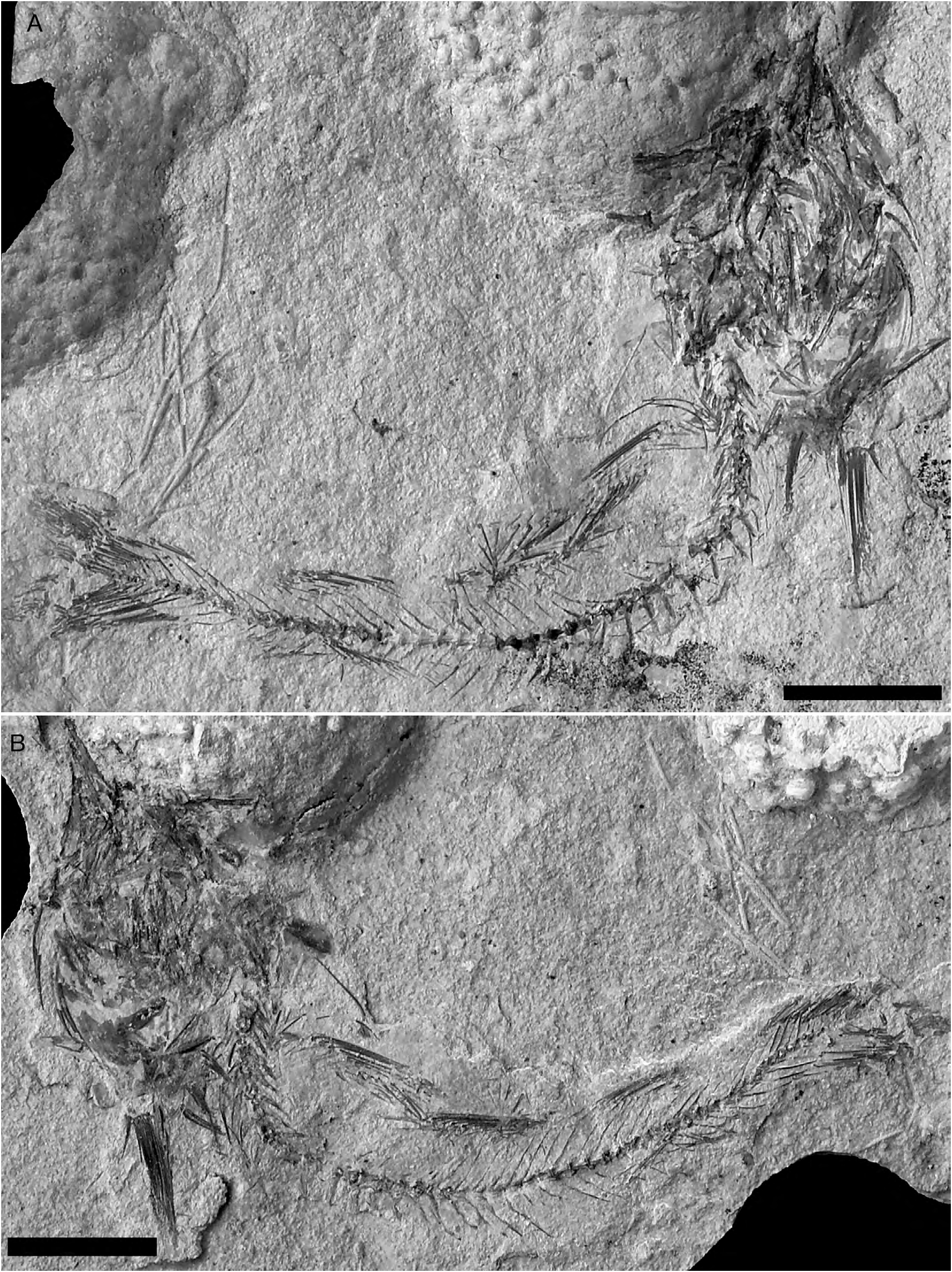
REFERRED SPECIMENS. — NHMW 1975/1752/248 and 247, complete well-preserved articulated skeleton in part and counterpart, 114.7 mm SL ( Figs 2 View FIG A-B); NHMW 1988/140/48a and b, nearly complete articulated skeleton ( Fig. 2C, D View FIG ); NHMW 1976/1812/38, partially complete articulated skeleton lacking the caudal portion ( Fig. 3A View FIG ); NHMW 1976/1812/51a and b, nearly complete articulated skeleton in part and counterpart ( Fig. 3B, C View FIG ); NHMW 2002z0181/0087a and b, nearly complete articulated skeleton in part and counterpart ( Fig. 4A, B View FIG ); NHMW 1975/1691/28a and b, complete articulated skeleton, 195 mm SL; NHMW 1974/1650/5, partially complete articulated skeleton; NHMW 1986/138/13a and b; NHMW 1975/1691/81, moderately well-preserved neurocranium exposed in ventral view; NHMW 1975/1691/27, largely incomplete articulated skeleton; NHMW 1975/1752/42, incomplete partially disarticulated skeleton; NHMW 1975/1752/50, partially complete articulated skeleton; NHMW 1975/1752/246, partially complete articulated skeleton; NHMW 1988/140/80a and b, nearly complete skeleton in part and counterpart; NHMW 1975/1691/89, largely incomplete poorly preserved articulated skeleton; NHMW 1975/1696/68, largely incomplete poorly preserved articulated skeleton; NHMW 1975/1752/220, incomplete articulated axial skeleton in part and counterpart; NHMW 1976/1812/62, incomplete articulated axial skeleton; NHMW 1976/1812/138, incomplete articulated axial skeleton; NHMW 1975/1696/53, largely incomplete articulated skeleton; NHMW 1975/1735/8, largely incomplete, partially disarticulated skeleton; NHMW 1975/1691/18, largely incomplete articulated skeleton; NHMW 1975/1696/52, moderately well-preserved articulated caudal portion of the skeleton; NHMW 1975/1812/69, largely incomplete partially disarticulated skeleton; NHMW 1975/1752/29, moderately well-preserved articulated caudal portion of the skeleton; NHMW 1975/1752/202, moderately well-preserved articulated caudal portion of the skeleton; NHMW 1976/1837/98, nearly complete poorly preserved articulated skeleton; NHMW 1988/140/142, incomplete articulated skeleton; NHMW 1975/1691/22, incomplete articulated skeleton lacking the head; NHMW 1976/1837/53, incomplete articulated skeleton; NHMW 1976/1837/185, largely incomplete poorly preserved articulated skeleton;
NHMW 1986/138/27, incomplete poorly preserved articulated skeleton; NHMW 1976/1837/189, poorly preserved articulated skeleton; NHMW 1992/151/11a and b, partially preserved head in part and counterpart; NHMW 1986/138/8, nearly complete poorly preserved articulated skeleton; NHMW 1986/138/31, incomplete articulated skeleton, lacking part of the head and the middle part of the body; NHMW 1975/1752/217, incomplete poorly preserved articulated skeleton; NHMW 1976/1812/40, slightly incomplete articulated skeleton; NHMW 2000z0135/0078, poorly preserved articulated skeleton in part and counterpart; NHMW 2002z0181/0088 + 0089, nearly complete neurocranium in part and counterpart; NHMW 2006z0220/0005, poorly preserved largely incomplete anterior part of the body; NHMW 2006z0220/0007, largely incomplete disarticulated anterior portion of the body.
TYPE LOCALITY AND HORIZON. — St. Margarethen locality, Kummer quarry; Middle Miocene, Late Badenian, Bulimina-Bolivina Zone, NN5b zone (see Schmid et al. 2001).
DIAGNOSIS. — As for the genus.
MEASUREMENTS. — (based on specimen NHMW 1975/1752/248; total length: 123.9 mm; standard length [SL]: 114.7 mm) as percentage of SL: maximum body depth: 20.3; head length: 29.3; head depth: 16.7; snout length: 11.0; caudal peduncle length: 6.3; caudal peduncle depth: 8.0; predorsal (1 st fin) distance: 37.5; predorsal (2nd fin) distance: 52.1; predorsal (3rd fin) distance: 71.7; preanal (1 st fin) distance: 46.0; preanal (2 nd fin) distance: 74.1; distance between 1 st and 2 nd dorsal fins:7.5; distance between 2nd and 3rd dorsal fins: 9.0; distance between 1 st and 2 nd anal fin:5.4; mouth gape extension:12.9; 1 st dorsalfin base length: 7.5; 2nd dorsal-fin base length: 10.3; 3rd dorsal-fin base length: 14.2; 1 st anal-fin base length: 23.7; 2nd anal-fin base length: 14.2; pectoral-fin length: 14.1.
DESCRIPTION
The body is elongate and laterally compressed. The head is well-developed and slender, contained slightly more that three times in SL. The snout is elongate. The diameter of the orbit cannot be measured in the examined material because of inadequate preservation; however, the orbit was probably rather large in origin. The caudal peduncle is moderately developed. The head skeleton is disarticulated, at least partially, in all the specimens examined; the bones are always displaced from their original position and are often extensively fragmented, so that is somewhat difficult to recognize the original morphology of the various elements.
Neurocranium
The neurocranium is often exposed in ventral view (see Fig. 5). It appears to be moderately depressed dorsoventrally, with a straight upper profile. The neurocranium is quite large; its maximum width measured in the postorbital sector is contained less than two times in the length of its base. In general, the neurocranium is nearly ovoid, with the length of its orbitorostral portion, measured at the level of the frontal sphenotic notch, slightly greater than the length of the posterior portion. The ethmoid bloc is strongly ossified. The vomer is large, massive and ventrally flattened, with a curved anterior margin; vomerine teeth are not preserved in the material examined. The lateral ethmoids have wide triangular lateral wings. The posterior process of the lateral ethmoid appears to be absent. The (fused) frontals are the largest bones of the skull roof. The ventral surface of these bones (see Fig.5) is strongly thickened and ornamented by longitudinal ridges and pits. The parasphenoid is robust, apparently straight, with a flat ventral surface. The basioccipital forms the floor of the posterior portion of the neurocranium. The topographic relationships between sphenotics, prootics, pterotics, and intercalars are identical to those of other gadine fishes (see e.g., Svetovidov 1948). The external surface of the prootics and intercalars is convex. The posterior corner of the pterotics consists of a thick and remarkably pointed spinous process. Nasal and infraorbital bones are inadequately preserved in the specimens examined.
Jaws
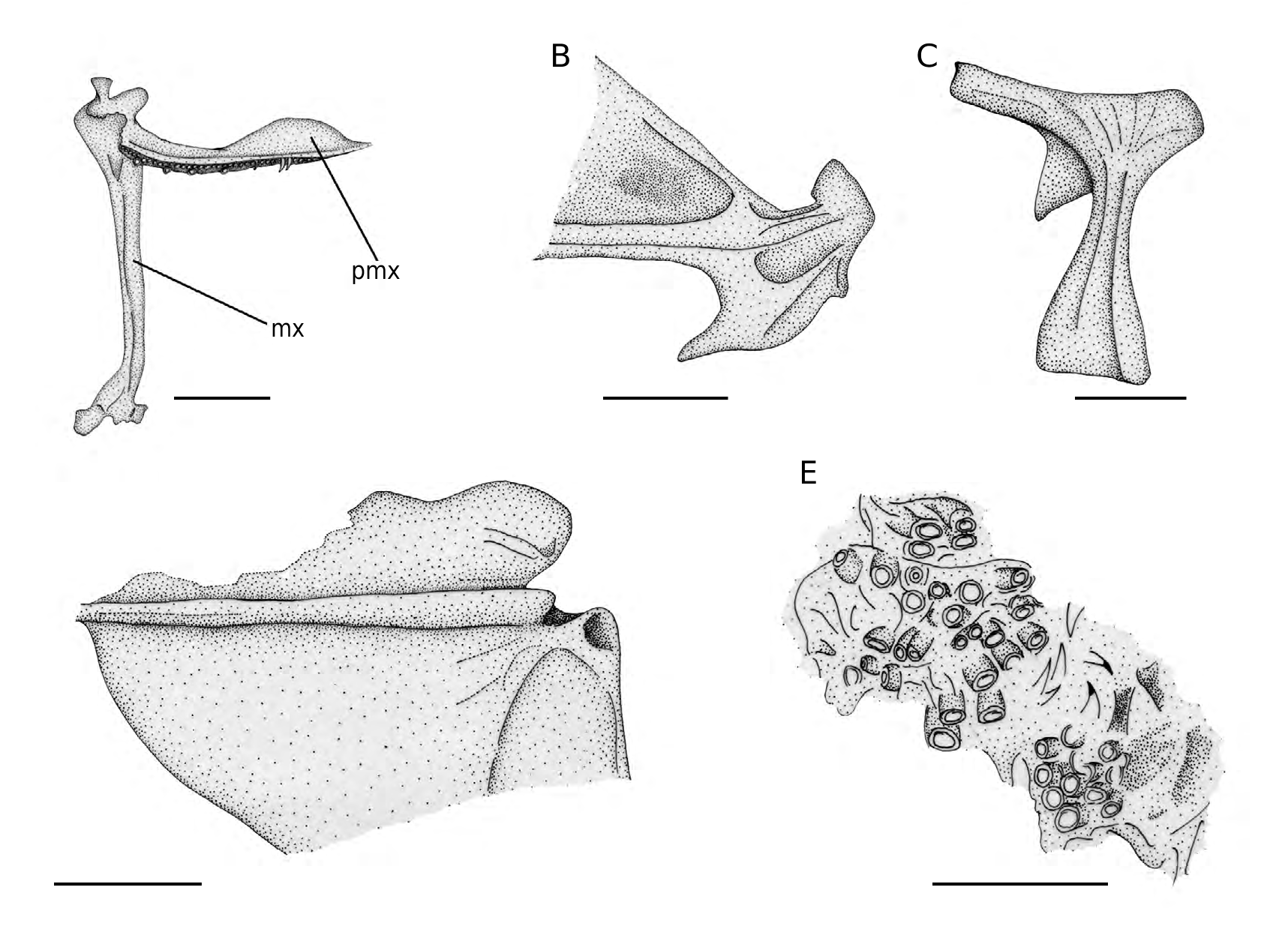
The gape of the mouth is rather wide; it probably reached the half of the orbit. The anterior tip of the dentary projects beyond the premaxilla. The premaxilla has robust and stout articular and ascending processes and an elongate and laminar postmaxillary process with gently rounded profile ( Fig. 6A View FIG ). There are two rows of teeth, the outer with few elongate and medially curved teeth, while the inner row is characterized by densely spaced minute pointed teeth. The maxilla is a robust and elongate bone, with a strong articular head ( Fig. 6A View FIG ). The lower jaw is long and shallow. The dentary has a long cavity (adductor chamber) that housed in origin the mandibularis section of the adductor mandibulae muscle. There is a short symphyseal process. The angulo-articular has a steep and irregular posterior slope and a large articular condyle ( Fig. 6B View FIG ). The mandibular teeth are similar to those of the upper jaw.
Suspensorium The bones of the suspensorium are extensively crushed and difficult to recognize, except for the fan-shaped quadrate and the hyomandibula; the hyomandibula ( Fig. 6C View FIG ) is relatively elongate and possesses a long and straight opercular process and a ventrally directed and sharply pointed preopercular process; the lower process of the hyomandibula is absent.
Opercular series
Opercular bones are badly damaged in most of the specimens examined, including NHMW 1974/1650/5, NHMW 1976/1812/69 and NHMW 1988/140/48. On the contrary, these are well preserved in the specimen NHMW 1975/1691/27. The preopercle is crescent-shaped with a regularly convex posterior margin. The interopercle and subopercle are laminar thin bones with a reduced thickening along their dorsal margin. The opercle has a subtriangular outline with rounded dorsal angles and shows a thick horizontal rib arising from its articular condyle ( Fig. 6D View FIG ).
Visceral arches
Of the branchial skeleton, fragments of elongate and slender bones, possibly the ceratobranchials, can be recognised in some specimens; curved and pointed (pharyngo) branchial teeth are also present and what appears to be a pharyngobranchial plate (2nd or 3rd) is preserved in NHMW 1976/1812/51b and NHMW 1986/138/13a, whereas the dentigerous subtriangular fifth ceratobranchials are exposed in NHMW 1976/1812/51 ( Fig. 6E View FIG ). The hyoid bar bears seven sabre-like brachiostegal rays, all articulating with the anterior ceratohyal.
Axial skeleton
The vertebral column comprises 45 to 46 vertebrae, of which 18 are abdominal.The vertebral centra are rectangular, longer than high, and characterized by lateral ridges. The anterior four vertebrae are shorter than the others. The abdominal vertebrae spo pto boc
FIG. 5. — Palimphemus anceps Kner, 1862 (NHMW 1976/1812/38), neurocranium, ventral view. Abbreviations: see Material and methods. Scale bar: 5 mm.
bear well-developed, nearly vertical parapophyses, characterized by broad bases and pointed tips. The neural spines of vertebrae two to 14 are thickened, antero-posteriorly enlarged, and strongly bent backward. Slender pleural ribs articulate with the abdominal vertebrae; the anterior ribs originate ventrally on the vertebral centra, while the posterior ones insert on the posteroventral margin of the parapophyses. Thin epineural bones are also present. The caudal skeleton (Fig. 7) is consistent with that of other gadids, and is characterized by two hypurals, a parhypural, two epurals, and absence of X and Y bones (see Barrington 1935a, b; Rosen & Patterson 1969; Marshall & Cohen 1973; Matarese et al. 1981; Patterson & Rosen 1989). The caudal fin is forked. The total number of caudal-fin rays ranges from 41 to 43, with seven upper and eight lower procurrent elements.
A
D
Median fins
There are three dorsal fins. The first dorsal fin inserts at the level of the 7th vertebra and contains 10-12 rays. The second dorsal fin inserts at the level of the 14th vertebra and contains 10-12 rays of progressively decreasing size. The first pterygiophore of the two anterior dorsal fins are large, roughly triangular in shape. The third dorsal fin originate at the level of the 26th vertebra and comprises 17-21 rays. The dorsal fins are separated from each other by large gaps, of which that between the second and third fins is greater than the length of the first dorsal-fin base. There is an indeterminate number of rayless pterygiophores (interneural bones in the sense of Dunn & Matarese 1984) between the second and third dorsal fins.
There are two anal fins. The first anal fin originates well behind the first dorsal-fin origin, approximately at the level of the 12th vertebra, and apparently consists of 18-21 rays, but the original number was probably higher. The length of the base of the first anal fin is widely shorter than the distance between the tip of the snout and the origin of the first anal fin. The second anal fin is approximately opposite to the third dorsal fin, inserting at the level of the 27th vertebra; this fin contains 18-19 rays. The first and second anal fins are separated by a short distance, which reaches about 5% SL. Despite their external separation, the two anal fins are internally continuous, connected by rayless pterygiophores.
Paired fins
The pectoral fin contains 15-18 rays. Of the girdle, the posttemporal, supracleithrum, cleithrum, postcleithrum, and parts of the scapula and coracoid are preserved in some specimens. The posttemporal is widely forked ( Fig. 8C), with the two arms form-
hyp3-5
phy
FIG. 7. — Palimphemus anceps Kner, 1862 (NHMW 1975/1691/28a+b), reconstruction of the caudal skeleton, left lateral view. Abbreviations: see Material and methods. Scale bar: 2 mm.
ing an acute angle that measures about 30°, very similar to that of Micromesistius australis Norman, 1937 (see Svetovidov 1948); the lateral flop (in the sense of Endo 2002) of the posttemporal is absent. The supracleithrum is a robust elongate bone with a gently rounded anterior profile. The cleithrum is large ( Fig. 8A), with a prominent lower arm, similar to that of other gadids (see e.g., Dunn 1989; Endo 2002). The postcleithrum is moderately elongate, slender and remarkably curved and possesses a laminar spatulate proximal head ( Fig. 8B); it greatly resembles that of M. australis (see Svetovidov 1948).
The pelvic fin consists of six rays. The basipterygium ( Fig. 8D) is narrow and tubular distally and broadly expanded proximally; the proximal portion of the bone is roughly triangular with a thick central ridge, and a pointed and nearly triangular median process.
Squamation
The body is covered by small cycloid scales.



| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Palimphemus anceps Kner, 1862
| Carnevale, Giorgio, Harzhauser, Mathias & Schultz, Ortwin 2012 |
Palimphemus anceps
| HOFMANN T. 2001: 221 |
| SCHMID H. P. & HARZHAUSER M. & KROH A. 2001: 22 |
| BRZOBOHATY R. & SCHULTZ O. 1978: 444 |
| SCHUBERT R. J. 1906: 697 |
| WOODWARD A. S. 1901: 270 |
| KNER R. 1862: 495 |