
Rhamdia enfurnada, Bichuette & Trajano, 2005
|
publication ID |
https://doi.org/10.1590/S1679-62252005000400016 |
|
DOI |
https://doi.org/10.5281/zenodo.15864921 |
|
persistent identifier |
https://treatment.plazi.org/id/631587CB-151B-DD5E-FED9-FDC3FA67F81F |
|
treatment provided by |
Carolina |
|
scientific name |
Rhamdia enfurnada |
| status |
sp. nov. |
Rhamdia enfurnada View in CoL , new species
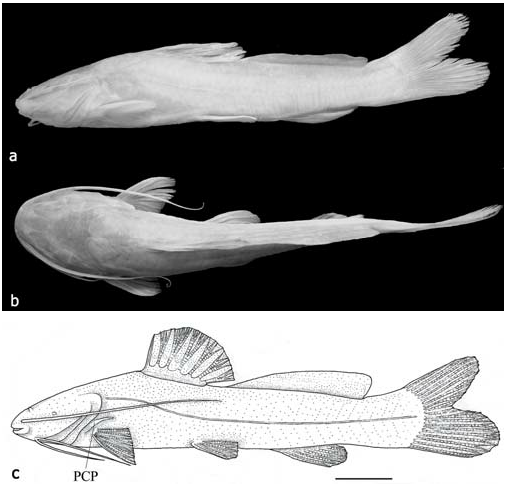
Fig. 1 View Fig
Holotype. MZUSP 87776, 137.8 mm SL, Brazil, southwestern State of Bahia, Serra do Ramalho, Município de Coribe , Povoado de Descoberto , Gruna do Enfurnado ( 13º38’44"S 44º12’05.9"W, elevation 628 m), middle São Francisco River basin; E. Trajano & M.E. Bichuette, 5 Sep 2003. GoogleMaps
Paratypes. MZUSP 87777 View Materials ( 13 exs.), 50.5-218.5 mm SL ; MZUSP 87778 View Materials ( 1 ex., c&s), 58.9 mm SL ; MZUSP 87779 View Materials ( 1 ex., c&s), 99.7 mm SL; and LIRP 5643 View Materials ( 4 exs.), 43.8-147.1 mm SL, all collected with the holotype .
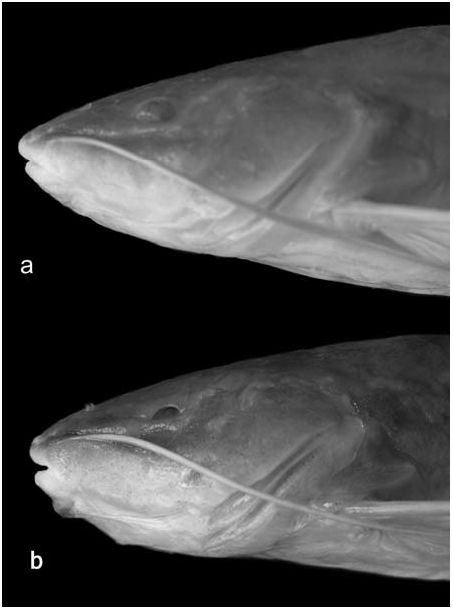
Diagnosis. Rhamdia enfurnada differs from its epigean congener ( Rhamdia cf. quelen ) from the São Francisco River basin by the highly variable eye size, ranging from reduced to externally invisible, with an open orbital concavity (orbital diameter varying from 6.4 to 13.6% of head length in R. enfurnada and from 14.5 to 24.4% in the epigean species); eyes, when present, pigmented and covered by semi-opaque skin, orbital rim absent or poorly developed in most specimens, well-defined only in some small specimens; dark pigmentation reduced due to a decrease in density of melanophores; and iridorophores present and concentrated in the opercular region. In comparison to other troglobitic Rhamdia species , R. enfurnada exhibits a lower degree of troglomorphy regarding eyes and pigmentation, which are highly variable; Rhamdia reddelli is described as lacking eyes and with almost no pigmentation; likewise, R. macuspanensis and R. laluchensis as having rudimentary eyes, covered with skin and not visible externally, and with reduced body pigmentation; Rhamdia zongolicensis is also almost devoid of dark pigmentation, but eyes are less reduced than in R. reddelli . Rhamdia quelen urichi , from Trinidad, also does not exhibit any trace of eyes externally, but the body coloration is described as pale yellowish brown. Rhamdia enfurnada has a concave dorsal profile of the head and a complete absence of pigmentation and eyes. Variability in development of eyes and pigmentation as in R. enfurnada was only reported for Rhamdia laticauda typhla , from Belize.
Description. Morphometric data in Table 1 View Table 1 . Distribution of standard length classes relative to 121 individuals measured under anesthesia and released in July 2005 during ecological study presented in Fig. 2 View Fig .
Body elongated, its greatest depth at vertical passing by dorsal-fin origin ( Fig. 1a View Fig ). Dorsal profile anterior to dorsal fin slightly to distinctively convex, then descending to caudal peduncle, slightly concave between dorsal and adipose fins. Head depressed, broad and relatively short, trapezoidal in dorsal profile. Largest specimen of R. enfurnada , 218.6 mm SL, shows signs of age, including emaciation and arched backbone, probably being close to maximum size of species ( Fig. 3 View Fig ).
Mouth terminal, upper jaw slightly in advance of lower jaw. Rictal folds well developed, reaching or slightly surpassing posterior margin of orbit. Premaxilla with single band of small, conic teeth in 10 irregular tooth rows, its posterolateral corners rounded, not produced. Dentition of lower jaw similar to that of premaxilla, in six irregular tooth rows. Palatine and vomer edentulous. Maxillary barbel long, reaching origin of anal fin in smaller specimens, and between middle and tip of depressed pelvic fin in larger individuals. Mental barbels long, inner-mental barbel surpasses posterior border of branchial membrane, almost reaching pectoral-fin insertion; outer-mental barbel reaching tip of depressed pectoral fin. Inner mentalbarbel base inserted slightly in advance of outer mental-barbel base. Anterior nares tubular, near border of snout; posterior nares tubular (incomplete in some specimens) and short, with rounded openings; internarial length less than width between posterior nares. Eye variable in size ( Fig. 4 View Fig ). Branchial membranes touching midventrally, but not overlapping (except in largest examined specimen, where membranes slightly overlap), and united to isthmus only anteriorly. Anus and urogenital papilla separated; urogenital papilla conspicuous and variable in size and shape, from short and triangular to elongate, located at midlength between bases of anal and pelvic fins.
Dorsal fin with i,I,7 rays. Dorsal spine weakly developed, smooth, without serrae, comparatively short, its length 39.6- 77.7% of dorsal-fin height.Adipose fin long (30.2-36.0% SL), its origin at vertical through middle of depressed pelvic fin, and its end slightly posterior to end of depressed anal fin. Adipose fin with slightly convex dorsal margin and conspicuous posterior free lobe. Caudal fin forked, ventral lobe slightly longer than dorsal; ventral lobe rounded in most individuals, pointed in some small specimens, dorsal lobe variable, pointed (in most small fish) to rounded. Dorsal lobe of caudal fin with i,7 and ventral lobe with i,8 principal rays. Anal fin with v,7-8 rays; its margin rounded in most specimens, pointed in some small fishes.
Pectoral fin with I,9 rays. Pectoral spine well developed, depressed, elliptical in transversal section, its anterior margin with 4-5 retrorse serrae distally and 7-13 small serrae proximally, posterior margin distinctly serrated ( Fig. 5 View Fig ). First branched pectoral-fin ray longest, other branched rays decreasing in length posteriorly. Posterior cleithral process relatively long, strong, reaching one-third of depressed pectoral fin ( Fig. 1c View Fig ). Pelvic fin with i,5 rays, its origin at vertical through end of dorsal-fin base, pointed in smaller specimens, rounded in larger ones.
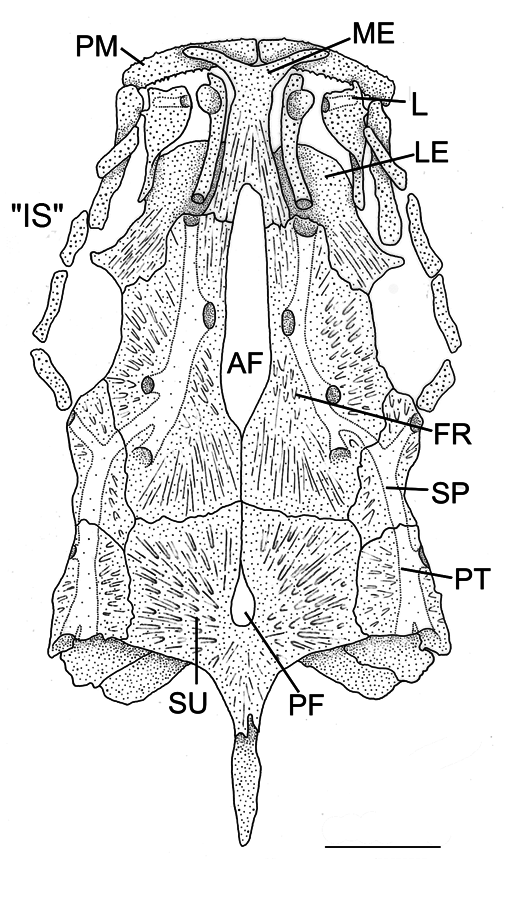
Skull roof ( Fig. 6 View Fig ) with wide, elongated anterior cranial fontanel; mesethmoid contacts frontal posteriorly on either side of concavity formed by anterior fontanel; frontal broad, greatest width equal to length, meeting posteriorly along 1/4 of its length. Posterior cranial fontanel reduced to elliptical foramen near center of supraoccipital. Supraoccipital process short, its length equal to length of supraoccipital bone excluding posterior process. Infraorbital bone series comprising smaller orbital cavity. Branchiostegal rays six.
In two cleared and stained specimens: 38 post-Weberian free vertebrae; nine pairs of ribs. Caudal skeleton with four hypural plates: rectangular parhypural; triangular hypurals 1+2; triangular hypurals 3+4, triangular hypural 5.
Color in alcohol. Specimens light gray to yellowish; base of dorsal fin, margin of adipose and dorsal fins slightly duskier, especially in darker fish; faint lateral stripe in darker fish, usually interrupted; melanophores concentrated on pseudotympanum in some specimens (not related to general degree of pigmentation); body coloration pattern otherwise homogeneous, pectoral, pelvic, anal, and caudal fins light colored. Melanophores concentrated on top of head and, in 11 out of 20 fish, also behind orbit, forming dark spot slightly larger than orbit. Density of melanophores in three cave and in three epigean specimens, respectively: 22.4, 28.8, and 30.4 melanophores per mm 2 in Rhamdia enfurnada , and 43.2, 46.4, and 51.2 melanophores per mm 2 in Rhamdia cf. quelen , from same river basin (MZUSP 54640).
Color in life. Light gray to yellowish, with melanophores concentrated on head region and margins of adipose and dorsal fins. Iridophores present, concentrated on opecular region, forming yellowish iridescent spot covering 2/3 of this region.
Distribution. Known only from its type locality, the Gruna do Enfurnado ( 13º38’44"S 44º12’05.9"W, elevation 628 m), Serra do Ramalho, Bahia State, middle São Francisco River basin, northeastern Brazil.
Etymology. From Gruna do Enfurnado, the name of the type locality, meaning shut, hidden, or isolated in a cave or cavern.
Habitat description. The Gruna do Enfurnado comprises 7,560 m of mapped passageways. This cave has about 3,000 m of subterranean streams and pools, with two main permanent reaches respectively at the proximal and distal ends of the cave. Flash floods occur in the rainy season, connecting these reaches; during the dry season, several pools of different sizes become isolated between them. The depth during the dry season varied from less than 0.2 m to 2-3 m in the large pools. Large amounts of organic matter are present. The fishes are especially abundant in the downstream reach, at the far end of the cave. Physico-chemical water variables measured with a digital Horiba apparatus in a deep pool at the proximal stream reach, respectively in September 2003 (end of the dry season) and July 2005 (middle of the dry season), were: temperature = 20.5 oC / 21.5 oC; pH = 7.47/ 8.25; conductivity = 0.597 ms cm-1 / 0.711 ms cm-1; dissolved oxygen (DO) = 0.39 mg l-1 / 0.54 mg l-1; salinity = 0.02% / 0.03%. The low values of DO measured in the dry season may have been due to the accumulation of organic matter associated to the slow movement of water. Several healthy catfishes were found in the pool, indicating a high tolerance to hypoxia in the species. The high pH and conductivity values are typical of karst waters. The average annual temperature in the region is 25ºC, thus the measured water temperatures were lower than expected since, due to the thermal isolation, the temperature in subterranean habitats usually follow the annual mean temperature.
Notes on ecology and behavior. Rhamdia catfishes were distributed throughout the cave, from shallow, pebble-bottomed reaches, where smaller fish concentrate and may attain densities of 10 individuals m-2, to deep, slow-moving, soft-bottomed pools, where we found the larger specimens. The largest numbers of fishes were found at the distal permanent stream reach (around 150 individuals observed along the 600 m reach in July 2005 - Trajano & Bichuette, unpubl. data). A mark-recapture study is in progress aiming to estimate the population size, minimal extension of catfish movements and life cycle parameters. Examination of the stomach contents of the two cleared and stained individuals revealed a diet of invertebrates, mainly arthropods, including terrestrial items (fragments of Diplopoda and Ensifera fallen accidentally or washed into the water) and a great amount of detritus.
The catfish behavior was directly observed in the cave habitat. In addition, seven specimens were brought alive to the laboratory and kept collectively in a 100-l aquarium in a dark room. Like other troglobitic siluriforms ( e. g., P. kronei , Taunayia sp. , the new heptapterid genus from Chapada Diamantina, and the undescribed Trichomycterus species from Bodoquena and Serra do Ramalho, respectively Trichomycterus sp. 2 and 3 in Trajano, 2003), Rhamdia enfurnada exhibited an increased midwater activity when compared to their typically bottom-dwelling epigean relatives. Hiding places made of limestone blocks were available; the catfish exhibited variable cryptobiotic behavior, with smaller individuals hiding less frequently than the larger ones. The reduction of cryptobiotic habits is another difference in relation to epigean relatives shared by several unrelated troglobitic siluriforms ( Trajano, 2003).
The catfishes from Gruna do Enfurnado reacted to sudden illumination with flashlights and noise with increased swimming activity, which indicates that these catfishes perceive these stimuli. In reaction to light experiments, using choice-chamber (as those described in Trajano & Gerhard, 1997, among others), these catfishes were mostly indifferent to light, i.e., they did not show significant preferences, either to the dark chamber or to the illuminated one (A.C. Barbosa, unpubl. data).
The catfishes from Gruna do Enfurnado were moderately aggressive under laboratory conditions: when two individuals met, apparently by chance (no spontaneous active pursuing was observed), usually the larger one chased away the smaller, in a quick pursuit, or the smaller fish rapidly avoided the other one, without pursuing. These interactions were usually observed during feeding with living Artemia , when increased activity enhanced the chances of meeting, and may be mostly related to competition for food. Nevertheless, as expected in view of the high population densities observed in the cave, in general the catfish seemed to be quite tolerant to the proximity of conspecifics, since no individual exhibited clear signals of stress, even after having been kept together for one and one-half years. Weak, size-related dominance was mostly displayed in relation to food and hidden places.



| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |