
Esopinae Guinot & Van Bakel, 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4766.1.5 |
|
publication LSID |
urn:lsid:zoobank.org:pub:0E43BB66-03FD-443E-9D6E-1BEE52B0C459 |
|
DOI |
https://doi.org/10.5281/zenodo.3803785 |
|
persistent identifier |
https://treatment.plazi.org/id/663987C6-FFAB-A60C-B6F8-FAAAFC60F923 |
|
treatment provided by |
Carolina |
|
scientific name |
Esopinae Guinot & Van Bakel |
| status |
subfam. nov. |
Subfamily Esopinae Guinot & Van Bakel View in CoL , subfam. nov.
Type genus. Esopus A. Milne-Edwards, 1875 ( type species by monotypy: Esopus crassus A. Milne-Edwards, 1875 ).
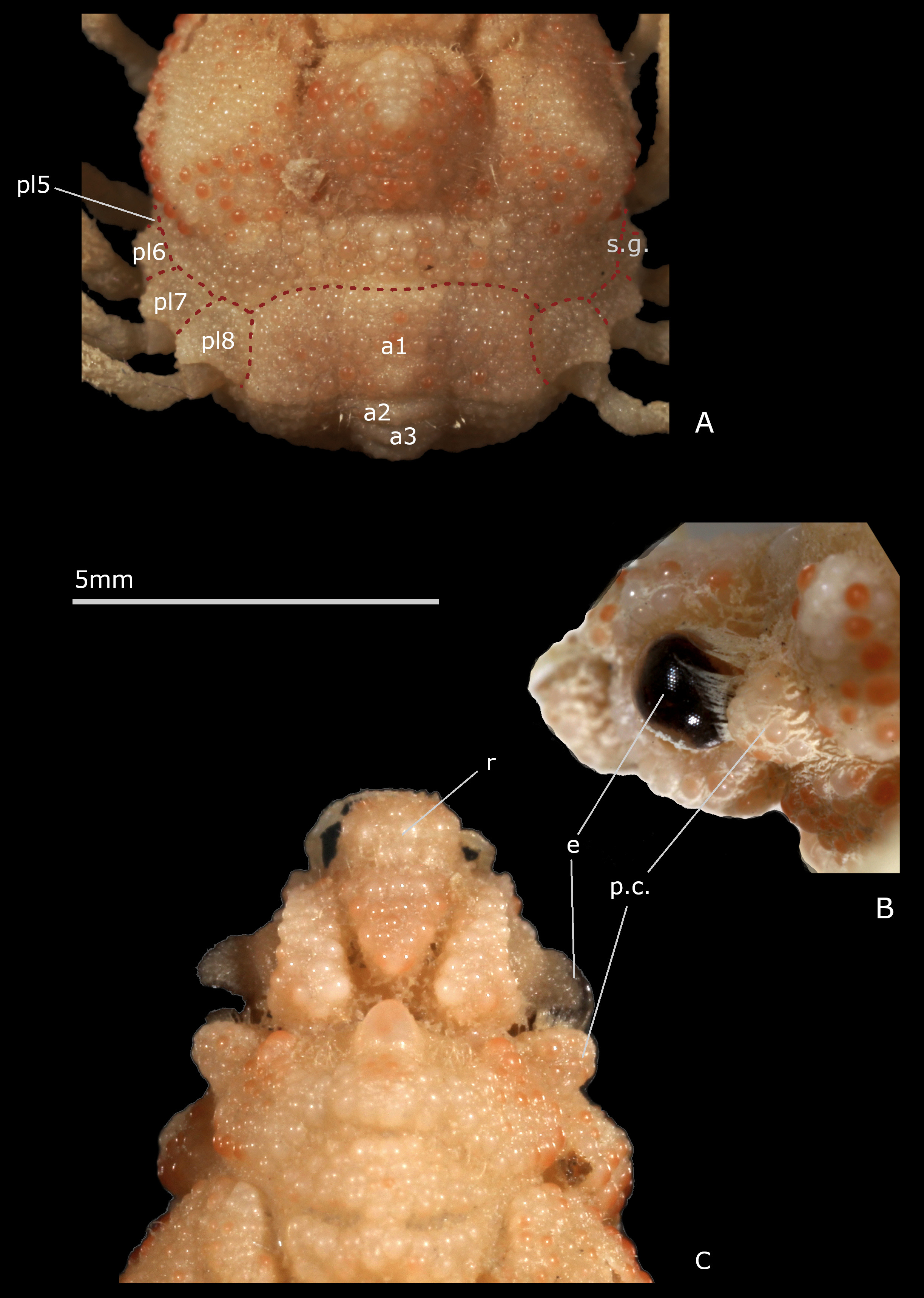
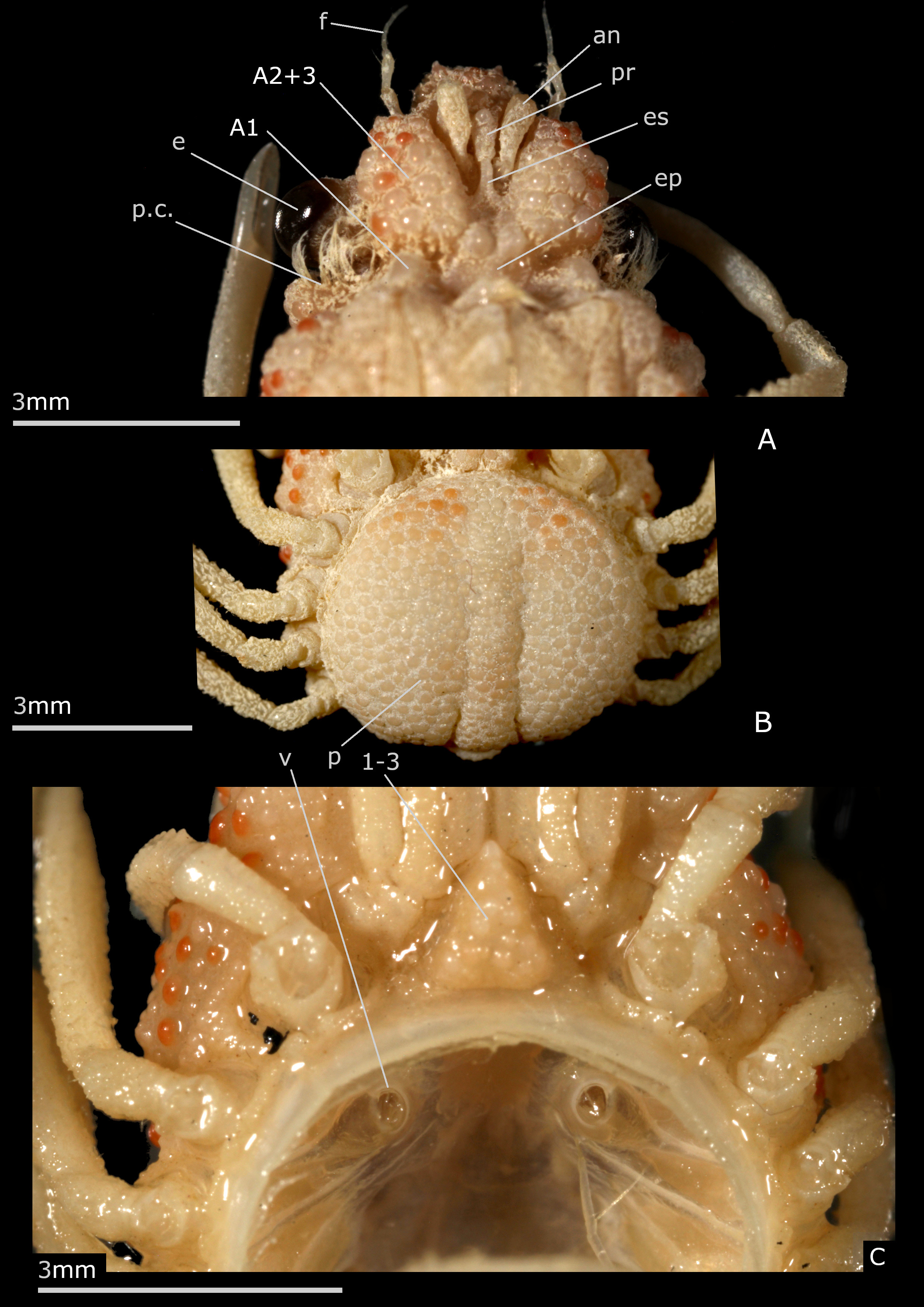
Description. Body thick. Carapace much longer than wide, narrow, gibbous, resting on setting gutter receiving carapace edge. Whole body with very thick cuticle. Dorsal surface sculpted, with inflated regions: frontal (tripartite), gastric (subdivided into several swollen portions), cardiac (prominent), intestinal (flat), delineated by several deep grooves. Exposure of latero-external portions of pleurites 5–8, calcified and ornamented like dorsal carapace surface, forming wide collar all around posterolateral margins of carapace; first (male and female) pleonal somite dorsal and similarly included in collar; pleonal somites 2–3 and portion of 4 dorsally visible. Portion in front of eyes delimited by deep depression and marked constriction; interorbital space divided into three nearly equal elevated lobes by two longitudinal depressions. Front broader than long. Rostrum wide, blunt, strongly folded downwards in the form of protruding, narrow, beak-shaped piece, positioned between antennules (as proepistome) and joining long anterior process of epistome. Antennular fossae (arthrodial cavities of antennules) very narrow, closed; antennules folded almost longitudinally. Antenna: articles 1 (urinary) and 2+3 (basal) adjacent; basal article coalescent with rostrum and epistome, very large, without keel, prominent, extending well beyond eyes, each expanding medially on epistome and thus close to the corresponding article of the other side; its distal and disto-lateral portions visible dorsally; other antennal articles proportionally very small in size; articles 4, 5 free; flagellum very short. Preorbital tooth absent. Ocular peduncle stout, protected in orbit. Postorbital tooth thick, cup-shaped to receive eye, fused with rounded subhepatic region. Epistome rather small. Buccal cavity wide. Milne-Edwards openings separated from chelipeds, entirely filled by developed mxp3 coxa. Mxp3 flat, completely covering buccal frame, operculiform; merus slightly dilated outward and forward, slightly notched on inner side for insertion of palpus; crista dentata with small teeth and row of setae. Branchiostegite extremely reduced. Male chelipeds very long, usually more than twice as long as the carapace; large gap between the fingers, near base. Female chelipeds shorter, less than length of carapace; fingers not gaping. Ambulatory legs very slender, rather long, unarmed; merus without spine on dorsoproximal margin; dactyli very thin, long, not subchelate, applying along propodi. Female sterno-pleonal cavity deeply hollowed. Female pleon wide, swollen, forming large disc limited by high sternal ridge, and formed of four somites, the somites 5, 6 being coalescent into pleotelson. Male pleon composed of somites 1–5 free plus pleotelson (somite 6 plus telson). Female pleon composed of somites 1–4 free plus pleotelson (somites 5, 6 plus telson). Male thoracic sternum markedly wide, with strong ridges and depressions, granular on exposed surface, smooth inside sterno-pleonal cavity; anterior shield formed by fused sternites 1–3 without demarcation in both sexes, as triangular, inflated, produced plate inserted between mxp3; a depression at level of suture 3/4; lateral extension of sternite 3 very short, hardly visible; sutures 4/5–7/8 interrupted; deep small depression at base of sternite 8, instead of median line. Sternum/pterygostome junction complete due to curved anterior extensions of sternite 4. Sternal extensions joining exposed pleurites (sternum/pleurites connections): wide between P1/P2, narrower between P2/P3, P3/P4, P4/P5. Female thoracic sternal sutures obliquely directed forward. Vulvae produced, on anteriorly displaced sternite 6. Partial condylar protection of penis, penis emerging from anterior border of P5 coxo-sternal condyle. G1 with broadened half proximal portion, and apical lobe bearing aperture; G2 very short. Pleonal-locking system consisting of buttons on sternite 5, sockets on pleotelson.
Remarks. In Esopus crassus the rostrum extends ventrally in a thick, beak-shaped piece, gradually tapering off, that is positioned between the antennules; it does not develop any spine ( Figs. 2B, C View FIGURE 2 , 3A View FIGURE 3 ); in oblique view, after removing the antennule, only a thick septum is visible. In Esopus it is the ventral part of the rostrum itself that just comes between the antennules and takes the place, at least without dissection of the region underneath, of the ‘proepistome’, the ‘antennular septum’ or ‘interantennular septum’ of authors. The proepistome is a structure defined as the “median, ventrally directed, cuticular outgrowth that separates the two cavities into which each antennule may be retracted” ( Ingle 1983: 6) or “the plate separating antennular cavities” (McLaughlin 1981: 167). In fact, the proepistome represents the sternite of the antennular somite (the first cephalic somite) and, if so, may connect to the epistome (sternite of the antennal somite) and thus attaches the carapace to the sternal surface. The term ‘outgrowth’ actually covers a set of modalities since in Brachyura this rostral outgrowth occurs in a wide variety of forms, and especially in Majoidea , in which the rostrum is often well developed and highly diverse. Among the Brachyura, the similarity of position (topographic and position in relation to other parts) of the rostral outgrowth is obvious, but a real homology between all the conditions is to be studied. In Esopus the last portion of the curved rostrum reaches the spiniform anterior process of the epistome. According to Cano (1893: 38, figs. 48a–c, 81) in spider crabs the primitive rostrum folds down to form a septum between the two antennules to join to a median process of the epistome; the interantennular septum would be the true rostrum of the zoea, the so-called rostrum of the adult being a secondary outgrowth of the front (see Balss 1929: 2; Garth 1958: 7–8). The different character states of the proepistome structure (often ignored by carcinologists) are very important for the phylogeny of Brachyura.
Another point deserves to be clarified. By using the term ‘fused’ loosely, it can mean that there is a real fusion process, thus without visible sutures (as in the case of the basal antennal article consisting of 2 and 3 fused articles in most Brachyura; or in the case of the thoracic sternum, where sternites are more or less fused); but it can also simply mean that two structures are in very tight contact but remain separate.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
SuperFamily |
Majoidea |
|
Family |