
Saccoglossus sonorensis, C. B. Cameron, C. Deland & T. H. Bullock, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.893894 |
|
DOI |
https://doi.org/10.5281/zenodo.6201833 |
|
persistent identifier |
https://treatment.plazi.org/id/68014E2E-FD35-FFB0-B8DD-FA78FE47FB80 |
|
treatment provided by |
Plazi |
|
scientific name |
Saccoglossus sonorensis |
| status |
sp. nov. |
Saccoglossus sonorensis View in CoL n. sp.
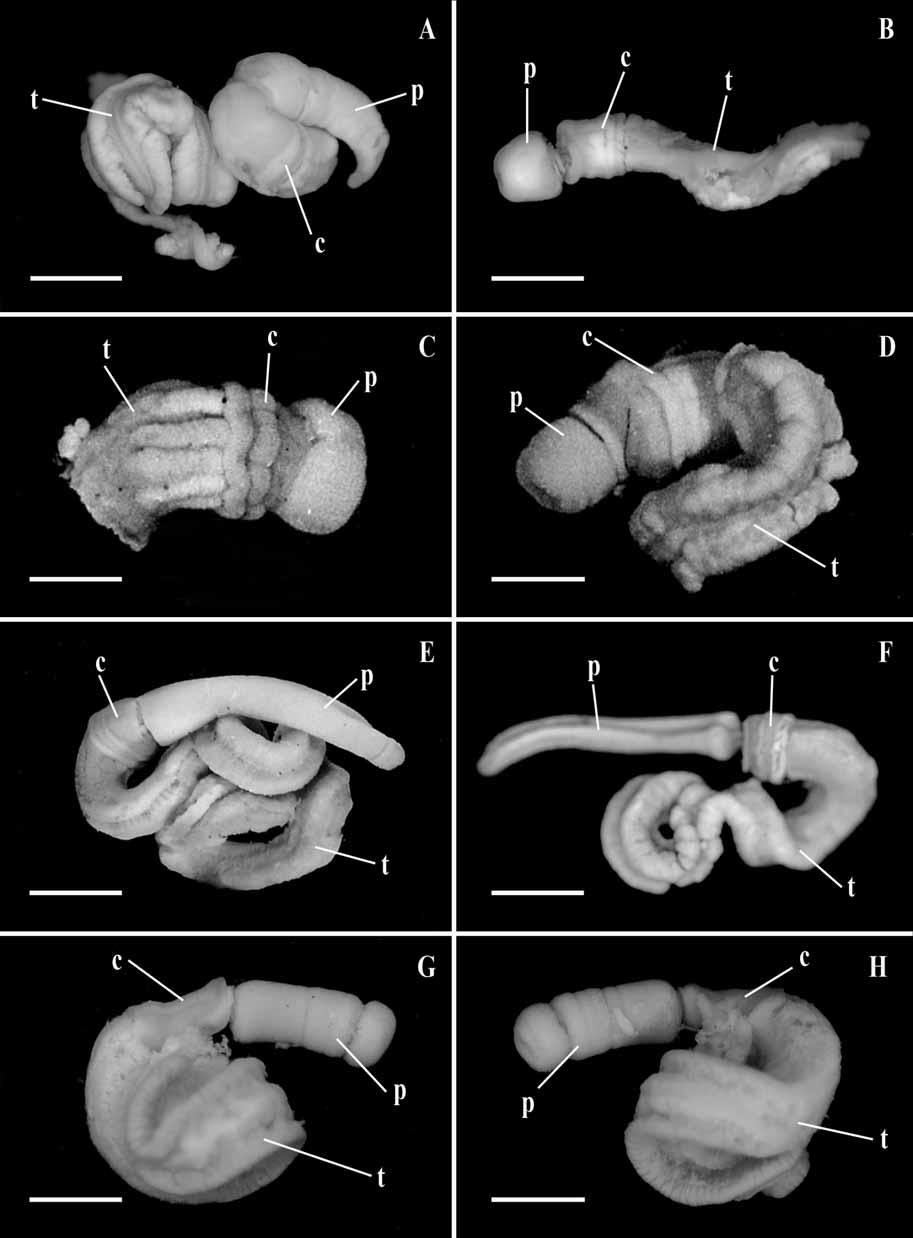
( Figs 1G, H View FIGURE 1 ; 5A–G)
Material examined. A single specimen in good condition which will be the type (T.H.B. accession no. 409) was collected from Cholla Bay, near Puerto Penasco, Sonora, Mexico (31° 17' N, 113° 35 W) in the Gulf of California, by T.H.B. in January, 1949, in slightly muddy sand. Three more anterior fragments were taken in January, 1957, at the same place and a dozen more in 1971. When an area with typical castings every few meters was found, nearly every casting yielding at least a tail fragment the habit similar to that of S. pusillus , S. kowalevskii , S. bromophenolosus and perhaps typical for the genus. Holotype: Accession no. NMNH 1132799. Paratype: Accession no. NMNH 1132800.
External features ( Figure 1 G, H View FIGURE 1 ): The total length is commonly 120-150 mm or more, the proboscis is 25 mm long, the collar is 2-4 mm. The proboscis is 2-3 mm wide and yellow orange; the collar is only slightly wider and a little deeper orange. Genital ridges are not conspicuous but the gonads commence immediately behind the collar. Ventral muscular ridges in the trunk are not present.
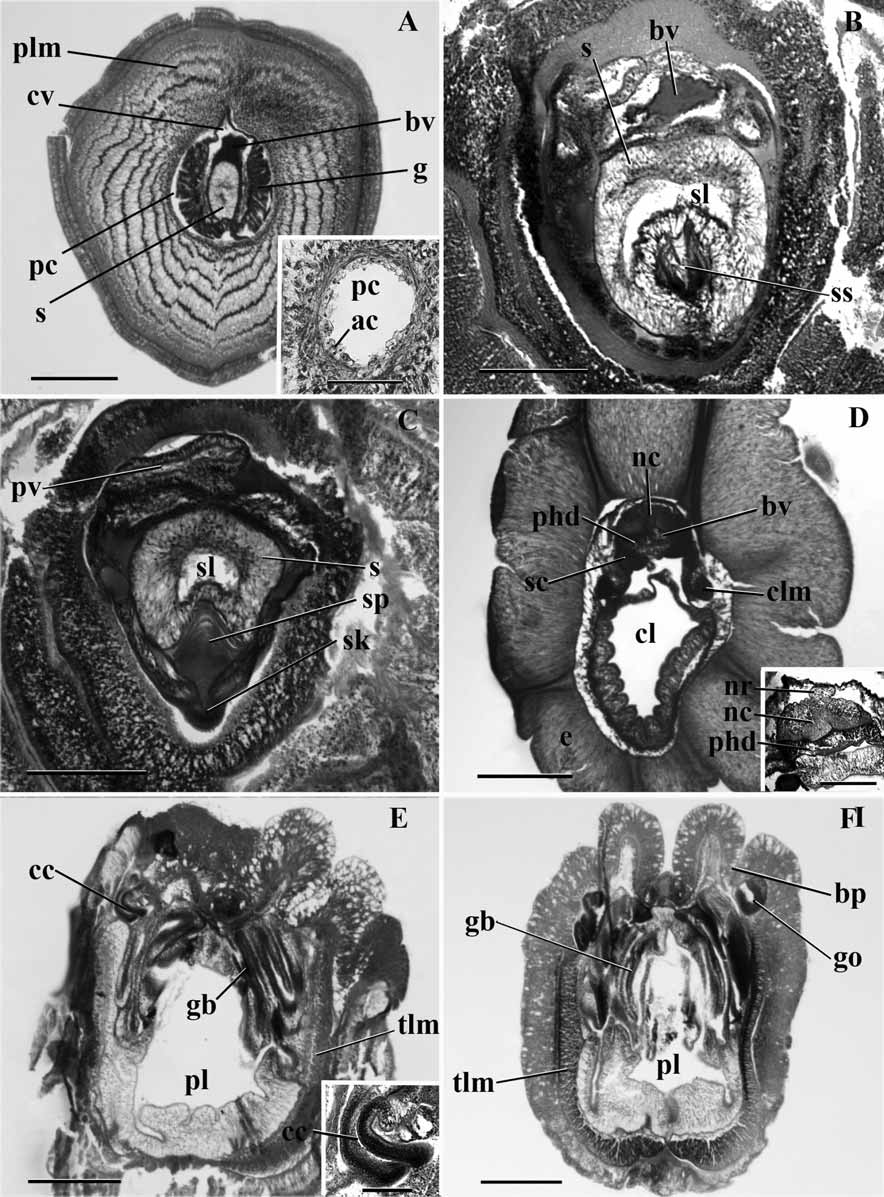
Internal features ( Figure 5 View FIGURE 5 ): The proboscis nerve fiber layer is rather thin, but shows a slight middorsal thickening. The circular muscle fiber layer is only about 4 fibers thick. Longitudinal muscle fibers are arranged in 5 to 6 concentric rings ( Fig. 5A View FIGURE 5 ). The proboscis lumen extends as a narrow round space almost to the tip of the proboscis, the anterior most part being occupied by connective tissue cells. It is lined by a layer of amoeboid cells in turn surrounded by a thin layer of circular fibers ( Fig. 5A View FIGURE 5 inset). The glomerulus extends anteriorly a little over the tip of the stomochord. The tip of the cardiac vesicle, which extends slightly beyond the stomochord, is surrounded on all sides by the glomerulus. The lumen of the stomochord extends to its tip only in the form of small irregular spaces, which are not continuous with each other. There is a dorsal glomerulus. The glomerulus covers even the ventral side of the stomochord, appearing in cross section as four or five finger-like processes hanging down from the ventral side of the stomochord. The cardiac vesicle is rather large and surrounds the stomochord on all sides except ventrally. But the lumen of the cardiac vesicle is quite filled with connective tissue, excepting the middorsal part ( Fig. 5A View FIGURE 5 ). The stomochord lumen has two ventro lateral caeca, equally well developed and continuous with the primary central lumen ( Fig. 5B View FIGURE 5 ). Posterior to these ventro lateral caeca, that is in the neck region, the wall of the stomochord becomes thinner and the stomochord now has a spacious and clear lumen. The end plate of the proboscis skeleton has a conspicuous middorsal spine which projects into the ventral caeca of the stomochord ( Fig. 5C View FIGURE 5 ). The proboscis skeleton has a distinct and deep keel over a short length in its anterior part ( Fig. 5C View FIGURE 5 ), but the keel is absent posterior to this, although the skeletal body is normal; the length of the keel antero-posteriorly is about 200 µm. Only the right proboscis canal is present ( Fig. 5C View FIGURE 5 ), occupying a median dorsal position posterior to the cardiac vesicle, and opening by a right proboscis pore. The dorsal and ventral septa are absent.
The collar epithelium is about 700 µm in thickness. There are no neuropores. The perihaemal cavities extend up to a few microns anterior to the proboscis pore. They are confluent at their anterior end. The collar nerve cord has neither a lumen nor any lacunae ( Fig. 5D View FIGURE 5 ). There are three dorsal roots arising from the collar nerve cord and entering the dorsal epidermal nerve layer. A dorsal mesentery is absent and the ventral one is confined to the posterior half of the collar. The skeletal cornua extend nearly to the posterior end of the collar, but just before their posterior extremity they take a sharp turn ventral wards from their dorsolateral position. The peribuccal diverticula are absent. The collar canals are vertically oriented ( Fig. 5E View FIGURE 5 , and inset). Their walls are not thick but the nuclei are arranged in several strata.
The openings of the branchial sacs on either side are in pairs, that is the left and the right pore are opposite to each other and not alternating. These openings are at the bottom of a deep and narrow groove along the sublateral line on either side of the middorsal line ( Fig. 5F View FIGURE 5 ). The gonads commence immediately behind the collar. They are simple, unlobed sacs which open individually onto the lateral (outer) wall of the sublateral groove. The gonads open along with the gill pouches and do not alternate with the latter. The branchial part is twice as large as the ventral part. The ventral and lateral aspects of the trunk coelom are filled with longitudinal muscle fibers but no ventro-lateral ridges are formed ( Fig. 5F View FIGURE 5 ).
Remarks. S. sonorensis does not resemble any known species of the genus in more than one or two of the usual specific characters. The only exception to this statement is S. otagoensis , which resembles the present species in the absence of the ventral muscular ridges in the trunk, the absence of the dorsal mesentery in the collar, the absence of the dorsal crest on the collar nerve cord and in the posterior extent of the skeletal cornua in the collar but differs in a number features such as the number of concentric muscle fiber rings in the proboscis, the external color, the side of the proboscis pore and the presence of a dorsal glomerulus.
Among the species of Saccoglossus commonly occurring on the west coast of America. S. pusillus resembles the present species only in the large branchial pharynx and in the posterior extent of the cornua. Other species of the genus like S. porochordus , S. rubur , S. kowalevskii , S. shumaginensis and S. rhabdorhynchus have at the most only one character each in common with the form consideration. Thus the specimen described above does not resemble any known species sufficiently well to warrant its inclusion in that species and hence a new species has been created to receive it. In the possession of only the right proboscis pore, it appears to be unique, but this character needs confirmation from several more specimens before it could be accepted as of specific value.
Etymology. The species is named after the Mexican state of Sonora.
Question marks indicate unknown character states. Three species are not included in this table: S. sulcatus ( Spengel, 1893) – only a figure available (no description), S. hwangtauensis ( Tchang and Koo, 1935) (unable to locate the paper), S. madrasensis ( Rao, 1957) (exists only as a conference abstract – there are no figures). S. cambrensis , S. serpentinus (Assheton, 1908) and S. ruber are synonymous species from the United Kingdom ( Burdon-Jones, 1960). There is little information available from the single specimen of S. ruber ( Tattersall, 1905) and so we used S. cambrensis (Brambell, Rogers and Goodhart, 1939) for our table.
Species characteristics were compiled from the following references: 1. Thomas, 1956; 2. Thomas, 1968; 3. King, Giray & Kornfield, 1994; 4. Brambell & Goodhart, 1941; 5. Kapelin, 1936; 6 & 7. Horst, 1939; 8. Benham, 1895; 9. Ritter, 1902, 1929; and 10. Brambell, Rogers & Cole, 1939; *New species.
Saccoglossus apatensis 1 Saccoglossus aulakoeis 2 Saccoglossus bromophenolosus 3 continued next page Saccoglossus horsti 4 Saccoglossus inhacensis 5 Saccoglossus kowalevskii 6
Proboscis shape? tapered anteriorly?
Proboscis length: width longer longer longer Proboscis longitudinal over 9 rings rings present but not well 4-5 rings musculature defined
continued next page Saccoglossus mereschkowskii 7 Saccoglossus otagoensis 8 Saccoglossus pusillus 9
Proboscis shape? cylindrical, rounded at tip continued next page continued next page Material examined. One specimen (anterior end through branchial region) was dredged at a depth of 250 m offshore NNW of Coos Bay entrance, OR (43° 21' N, 124° 21 W) by J. McNab on August 6th 1958 . Two fragments have also been dredged at a depth of 25 meters on July 30th 1958. Holotype: Accession no. NMNH 1132801.
External features: The proboscis is long and conical with a small groove in the posterior third of its length. The total length is estimated to be 1 cm. The length of the proboscis is 3 mm and the collar is 0.5 mm. In live material, the proboscis is white and the collar and trunk are salmon pink.
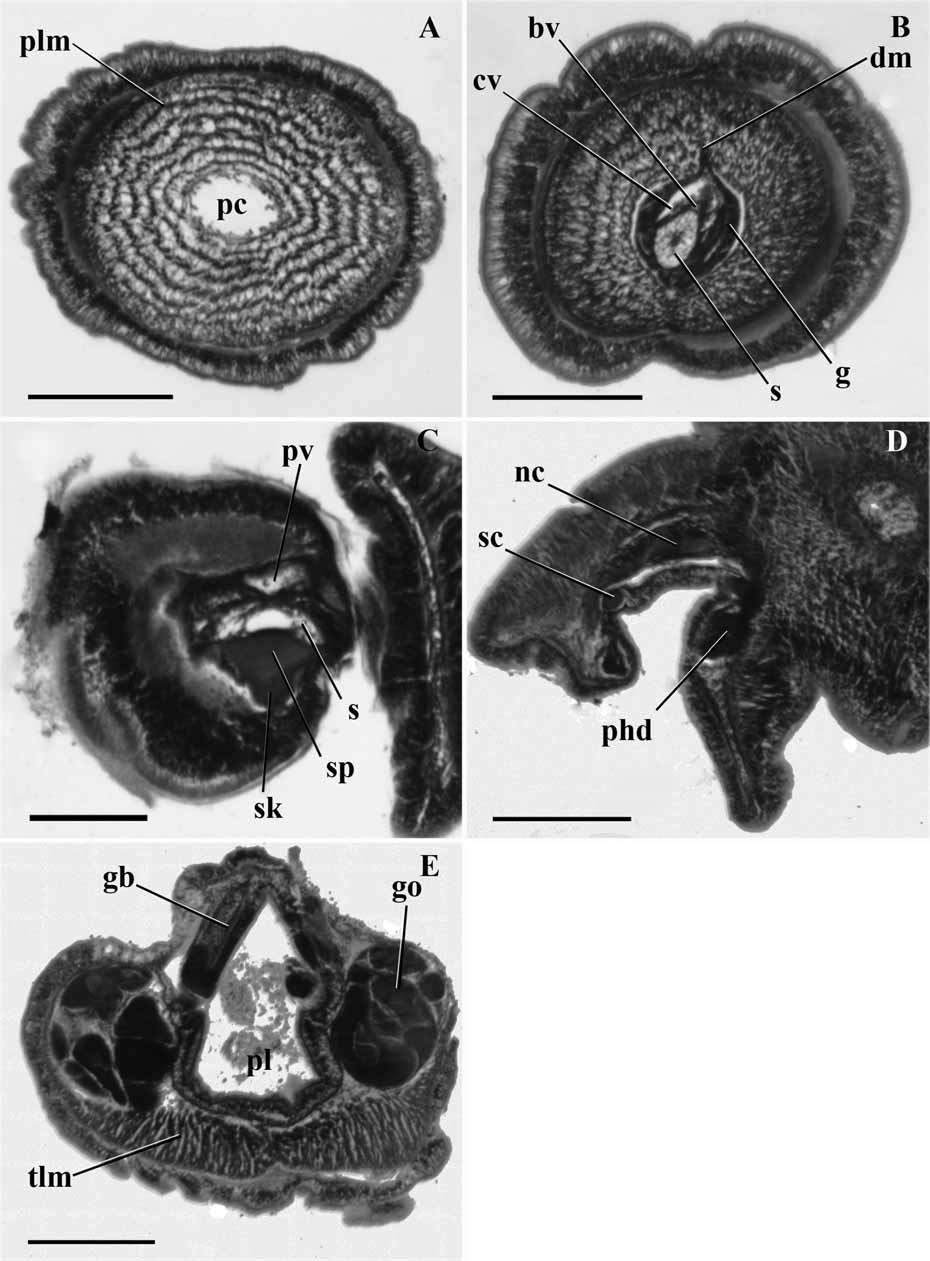
Internal features ( Figure 6 View FIGURE 6 ): The nerve fiber layer of the proboscis does not show any thickening along the dorsal midline. The proboscis coelom almost extends to the tip of the organ as a narrow cavity closely surrounding the proboscis complex ( Fig. 6A View FIGURE 6 ). The ventral and dorsal septa are present and start in the posterior half of the proboscis. They are both continuous throughout the length of the organ. The circular muscle fiber layer is thinner than the nerve fiber layer. The longitudinal muscle fibers are arranged in about 6 concentric rings ( Fig. 6B View FIGURE 6 ). The glomerulus extends over the tip of the stomochord and surrounds the entire proboscis complex. The cardiac vesicle is well developed and extends to the tip of stomochord. Its anterior half is filled with connective tissue. The stomochord has thick walls and a narrow lumen throughout its length. Its lumen presents a large ventral diverticulum, which is twice as big as the primary lumen and join each other at the beginning of the skeleton plate. The skeletal plate starts on the ventral side of the stomochord and the keel is obtuse and short. The two coelomic cavities in the neck are present and there is also one conspicuous left vesicle leading to the exterior by a left proboscis pore ( Fig. 6C View FIGURE 6 ).
Very few characters could be determined from the collar ( Fig. 6D View FIGURE 6 ) and trunk ( Fig. 6E View FIGURE 6 ) due to distortions in the sections. Only the few characters described below could be assessed. The dorsal mesentery of the collar is present, but it has been impossible to determine its extent, as for the presence and extent of the ventral one. An anterior neuropore may be present.
The branchial portion of the pharynx is equal in size to the ventral portion or slightly larger. There is a pair of dorso-lateral gonads, which are not lobed. The ventro-lateral longitudinal muscles of the trunk are well developed ( Fig. 6E View FIGURE 6 ).
Etymology. We dedicate the name of this animal to Professor A. Richard Palmer, University of Alberta, who has greatly contributed to our understanding of the comparative morphology and development of Pacific Coast invertebrates.
| NMNH |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.