
Otomops harrisoni, Ralph, Taryn M. C., Richards, Leigh R., Taylor, Peter J., Napier, Melanie C. & Lamb, Jennifer M., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4057.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:59548C19-0DE6-40BA-9956-45484E2EFB95 |
|
DOI |
https://doi.org/10.5281/zenodo.3860387 |
|
persistent identifier |
https://treatment.plazi.org/id/6A61B42F-FF8D-FF8A-84D3-FB3EFAEDF841 |
|
treatment provided by |
Plazi |
|
scientific name |
Otomops harrisoni |
| status |
sp. nov. |
Otomops harrisoni View in CoL sp. nov.
Harrison’s large-eared giant mastiff bat Figures 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10
Synonymy
Otomops martiensseni martiensseni: Harrison, 1965:2 View in CoL (part) Otomops martiensseni martiensseni: Hill and Morris, 1971:46 View in CoL Otomops martiensseni: Ðulic and Mutere, 1973:62 View in CoL (part) Otomops martiensseni: Kinoti, 1973:129 View in CoL
Otomops martiensseni: Mutere, 1973:83 View in CoL
Otomops martiensseni: Epelu-Opio, 1974:229 View in CoL
Otomops martiensseni martiensseni: Largen et al., 1974:250 View in CoL Otomops martiensseni: Kingdon, 1974:338 View in CoL (part)
Otomops martiensseni: Warner et al., 1974:171 View in CoL (part) Otomops martiensseni: Kayanja and Mutere, 1975:166 View in CoL Otomops martiensseni: Kayanja and Mutere, 1978:245 View in CoL Otomops martiensseni: Valdivieso et al., 1979:6 View in CoL
Otomops martiensseni: Freeman, 1981:61 View in CoL (part)
Otomops martiensseni: Norberg, 1981:365 View in CoL (part)
Otomops martiensseni martiensseni: Aggundey and Schlitter, 1984:144 View in CoL Otomops martiensseni: Fenton and Crerar, 1984:398 View in CoL Otomops martiensseni: Freeman, 1984:400 View in CoL (part)
Otomops martiensseni: Hickey and Fenton, 1987:381 View in CoL Otomops martiensseni: Norberg, 1987:53 View in CoL (part)
Otomops martiensseni: Norberg and Rayner, 1987 View in CoL :plate 1 (part)
Otomops martiensseni: Thollesson and Norberg, 1991:26 View in CoL (part) Otomops martiensseni: Long, 1995:1 View in CoL (part)
Otomops martiensseni: Peterson et al., 1995:178 View in CoL (part) Otomops martiensseni: Yalden et al., 1996:91 View in CoL (part)
Otomops martiensseni: Rydell and Yalden, 1997:72 View in CoL (part) Otomops martiensseni: Al-Jumaily, 1999:241 View in CoL
Otomops martiensseni: Monath, 1999 View in CoL :S130 (part)
Otomops martiensseni: Pearch et al., 2001:388 View in CoL
Otomops martiensseni: Debaeremaeker and Fenton, 2003:221 View in CoL (part) Otomops martiensseni: Jones and Rydell, 2003:303 View in CoL (part) Otomops martiensseni: Kock and Zinner, 2004:3 View in CoL
Otomops martiensseni: Kock et al., 2005:2 View in CoL
Otomops martiensseni: Taylor et al., 2005:26 View in CoL (part)
Otomops martiensseni: Lamb et al., 2006:46 View in CoL (part)
Otomops martiensseni: Lamb et al., 2008:25 View in CoL (part)
Otomops martiensseni: Tong et al., 2009:483 View in CoL (part)
Otomops martiensseni: Benda et al., 2011:25 View in CoL (part)
Otomops martiensseni: Lamb et al., 2012:8 View in CoL (part)
Otomops martiensseni: Patterson and Webala, 2012:5 View in CoL (part) Otomops martiensseni: Richards et al., 2012:913 View in CoL (part) Otomops martiensseni: Taylor et al., 2012:56 View in CoL (part)
Otomops martiensseni: Kading et al., 2013:2394 View in CoL (part) Otomops martiensseni: Ralph and Lamb, 2013:4234 View in CoL (part) Otomops martiensseni: Tao et al., 2013:739 View in CoL (part)
Otomops martiensseni: Yalden and Happold, 2013:480 View in CoL (part) Otomops martiensseni: Conrardy et al., 2014:259 View in CoL (part) Otomops martiensseni: Kassahun et al., 2015:168 View in CoL (part) Otomops martiensseni: Mortlock et al., 2015:1841 View in CoL (part)
Holotype. HZM 60.36217 (field number A51) is part of a series of specimens collected by Paul J. J. Bates, M. J. Pearch and O. Nurhussein on 19 July 1998. This is an adult male presently preserved in 70% alcohol. The cranium and baculum have been extracted and prepared. External and craniodental measurements are presented in Tables 1 and 2 View TABLE 2 , respectively. The cranium and mandible of the holotype are in good condition and are presented in figure 8. The baculum of the holotype was prepared following the methods of Hill & Harrison (1987) and Kearney et al. (2002), with slight modifications, and is illustrated in figure 9. This specimen was included in the morphological and mitochondrial DNA sequence-based analyses.
Type locality. Ethiopia, Bale District, Sof Omar Cave 06°54´N; 40°48´E; elevation 1340 m.
Paratypes. Thirteen adult specimens were collected from the same locality and on the same date as the holotype (n = 3 males, HZM 40.31315, HZM 44.31328, HZM 64.36221; n = 10 females, HZM 41.31316, HZM 42.31317, HZM 43.31318, HZM 56.36213, HZM 57.36214, HZM 58.36215, HZM 59.36216, HZM 61.36218, HZM 62.36219 (also DM 14750), HZM 63.36220). Measurements are provided in Tables 1–2.
Referred Specimens. Specimens originating from Djibouti, Eritrea, Ethiopia, Kenya and Yemen have been assigned to this taxon, as supported by molecular DNA and/or cranial morphological datasets, or based upon geographical location. Those specimens in bold have not been sequenced and their assignment to O. harrisoni is based on cranial morphological analyses alone. Djibouti: BMNH 69.1256, Day Forest National Park, Mount Day (11.767° N, 42.650° E, altitude 1400 m); Ethiopia: HZM 45.31369, HZM 46.31370, HZM 47.31371, HZM 48.31372, HZM 49.33964, HZM 50.33965, SMF 41832, SMF 41833, Bale District, Sof Omar Cave (6.900° N, 40.859° E, altitude 1300 m); Kenya: ROM 48654–48657, ROM 48659–48664, ROM 48666–48667, ROM 63772, ROM 63779, ROM 63782, ROM 63808, 19 km W of Makindu (2.300° S, 37.677° E, altitude 1000 m); MRAC 38546–38549, SMNS 46077, SMNS 46079, Chyulu Hills (2.583° S, 37.833° E, altitude 1930 m); ROM 65875–65879, Ithundu Caves, Kiboko (2.199° S, 37.717° E, altitude 920 m); ROM 81198–81199, Ithundu Caves, Makindu (2.333° S, 37.699° E, altitude 1100 m); ROM 68360, ROM 68362, ROM 68364, ROM 68366, Lake Baringo, Kampi Ya Moto (0.183° N, 35.867° E, altitude 1200 m); MRAC 35264, Machakos District (1.517° S, 37.267° E, altitude 1630 m); ROM 78155–78158, Makindu Cave, Makindu (2.300° S, 37.833° E, altitude 1000 m); ROM 65873, Makindu River (precise locality not defined); ROM 36517, ROM 36519, ROM 41920, ROM 41924, ROM 41927–41928, ROM 41932, ROM 78147–78148, ROM 78151–78152, ROM 78154, ROM 91249– 91250, Mount Suswa (1.150° S, 36.350° E, altitude 1895 m); ROM 79677, Nairobi (1.280° S, 36.817° E, altitude 1685 m); Yemen: HZM 39.31195, HZM 51.33976, HZM 52.33977, HZM 53.33978, HZM 54.33979, HZM 55.33980, NMP 91811–91816, SMF 87648–87649, SMF 87650, Hud Sawa Cave, Ar-Rayadi Al-Gharbi Mountains, 3 km NW of Al-Mahweet (15.483° N, 43.533° E, altitude 2150 m).
Etymology. This species is named after the late renowned mammalogist, taxonomist and bat expert Dr. David L. Harrison (1926–2015). Harrison’s numerous publications on Afro-Arabian Chiroptera , in particular the Molossidae , have significantly improved our knowledge of this poorly known family.
Diagnosis. Diagnosis is based on statistically supported morphological differences and the occurrence of strongly supported reciprocally monophyletic lineages observed in analyses of concatenated mitochondrial cytochrome b and D-loop DNA sequences. Three lineages were strongly-supported within the Afro-Malagasy region; O. madagascariensis was sister to the reciprocally-monophyletic sister lineages, O. martiensseni s.s. and O. harrisoni ( Fig. 2 View FIGURE 2 ). Otomops harrisoni is separated from O. martiensseni s.s., its sister lineage, by a genetic pdistance of 2.10%, and from O. madagascariensis by 3.40%. Otomops harrisoni is further diagnosed by unique molecular characteristics, i.e. strict synapomorphies in both the cytochrome b gene and the D-loop region ( Table 4 View TABLE 4 ).
In general, O. harrisoni is the largest and most robust member of the genus Otomops Thomas, 1913 , overlapping in certain external and craniodental measurements with O. martiensseni s.s. The pelage is short and velvety, appearing dark chocolate brown on the upper parts, with a distinct cream-coloured collar that extends from the nape of the neck to the back. The ventral pelage consists of lighter brown-coloured hairs; it extends to the wings and covers a section of the plagiopatagium. The borders of the body and wing membranes of both the dorsal and ventral surfaces are marked by a distinct yet thin band of white fur, extending from the mid-humerus to the upper reaches of the thigh. Ears are long (35.2–46.0 mm for males, 33.6–41.0 mm for females), forward-directed and are united by a flap of skin across the nose. The species is sexually dimorphic, with males characterised by an average forearm length of 70.7 ± 1.91 mm (64.0–74.0 mm, n = 41), whereas in females the average forearm length is 69.5 ± 1.62 mm (65.7–72.7 mm, n = 38).
In general, O. harrisoni can be distinguished from other Afro-Malagasy taxa by its long cranium and notably high braincase ( Fig. 8 View FIGURE 8 ). Otomops harrisoni is characterised by an average greatest skull length of 28.5 ± 0.36 mm (27.8–29.3 mm, n = 38) and 27.0 ± 0.36 mm (26.2–27.9 mm, n = 41), for males and females, respectively. The height of the braincase (excl. the tympanic bullae) of males averages 9.4 ± 0.18 mm (9.0– 9.8 mm, n = 38) and females average 9.1 ± 0.17 mm (8.7–9.5 mm, n = 41). There is distinct and comparatively deep depression across the fronto-parietal area of the braincase. The braincase is markedly domed in the frontal region. The lambdoidal crests are moderately developed, join to form a “V-pattern”, and extend to the sagittal crest at the highest point of the cranium. The sagittal crest remains slightly pronounced along the fronto-parietals, terminating at the posterior edge of a depression in the inter-orbital region. The rostrum is broad and robust, with fairly large, laterally flared nasals. The jugal process of the zygomatic arch is thickened and the zygomaxillary junction projects outwards. The external tympanic bullae are elongate and posteriorly broadened; they extend to the pterygoids and occupy a third of the braincase. The baculum of the holotype of O. harrisoni measures 1.16 mm ( Fig. 9 View FIGURE 9 ). The maximum width of the baculum shaft (SW) is 0.29 mm. The inflection of the baculum shaft (IB) in both the dorsal and ventral views is 54.53°.
The first nucleotide gives the ancestral state, followed by the nucleotide position in each sequence, and the last
nucleotide gives the derived state.
......continued on the next page Description and comparisons. Externals. Otomops harrisoni bears all the external morphological diagnostic features of the genus as described by Thomas (1913) and subsequently refined by Freeman (1981). As with other Otomops spp. the pelage is conspicuous and is characterised by dark chocolate brown fur on the back and head, with slightly paler under parts. A cream-coloured collar extends from the dorsal surface of the neck to the throat, separating the head and back regions. This collar varies in size amongst the members of the genus. It is fairly pronounced in the Afro-Malagasy members ( Monadjem et al. 2010; Goodman & Raherilalao 2014), and in some Indo-Australasian taxa ( Thomas 1913; Kitchener et al. 1992), yet is reduced to a narrow pale band of hairs in O. papuensis ( Lawrence 1948; Hill 1983). Characteristic of the genus, the large ears of O. harrisoni project forwards and join anteriorly above the nose. A series of brown spines is located along the anterior border of the ears (c. 15) and a flexible flange extends from the dorsal to the ventral surface of the pinnae; features shared by all Otomops spp. Similar to congenerics, the antitragus is absent and the tragus is minute. The two nostrils are stiffened by cutaneous ridges that run above and between them. These ridges are lined with small brown spines. The lips are characterised by loose folds and the anterior surface of the upper and lower lips are covered in short, light-brown bristles of hair. Common to other Afro-Malagasy and Indo-Australasian members, the upper lip projects beyond the lower lip. The feet of O. harrisoni are broad, with a comb of long white hairs. The ventral surfaces of the first and fifth toes are speckled with short light-coloured hairs. Adult males possess a distinct gular gland that is located on the lower portion of the throat. A similar gland has been reported from O. martiensseni s.s. ( Chubb 1917), O. madagascariensis ( Eger & Mitchell 2003) , and O. wroughtoni ( Thomas 1913) . The first digit of O. harrisoni is noticeably thickened at its base and tapers to a point at the claw. Some individuals possess thumb pads that are slightly distended. A series of O. martiensseni s.s. examined from Durban displayed similar thumb pads; this feature appeared to be most pronounced in males. Based on external measurements, O. harrisoni is, in general, larger than other Otomops spp., including Afro-Malagasy congenerics. For instance, the average forearm length in O. harrisoni is 70.7 ± 1.91 mm (63.8–74.0 mm, n = 41 males) and 69.5 ± 1.62 mm (65.7–72.7 mm, n = 38 females), whereas in O. martiensseni s.s. this measurement is 65.8 ± 1.60 mm (60.5–68.0 mm, n = 24 males) and 63.5 ± 1.53 mm (60.0– 66.3 mm, n = 22 females), and in O. madagascariensis average forearm length is 63.2 ± 1.35 mm (60.0– 66.0 mm, n = 20 males) and 61.2 ± 1.32 mm (60.0–64.0 mm, n = 18 females).
Craniodental characters. The cranium of O. harrisoni is, in general, longer and broader relative to other Afro- Malagasy Otomops spp., yet there is some overlap in measurements with other Afro-Malagasy taxa, particularly O. martiensseni s.s. ( Table 2 View TABLE 2 ). Given the morphological similarity between the two sister species, we have provided a side-by-side comparison of the crania of the holotypes of O. harrisoni and O. martiensseni s.s. to highlight the distinguishing features of the new species ( Fig. 10 View FIGURE 10 ). The average greatest skull length of O. harrisoni (males: 28.5 mm, range 27.8–29.3 mm; females: 27.1 mm, range 26.2–27.9 mm) is at least 1.04 times greater than the average greatest skull length of O. martiensseni s.s. (males: 27.5 mm, range 26.5–28.8 mm; females: 25.5 mm, range 24.7– 26.1 mm), and 1.12 times that of O. madagascariensi s (males: 25.7 mm, range 24.9–26.5 mm; females: 22.8 mm, range 22.8–24.5 mm). Multivariate analysis of craniodental data, demonstrated that O. harrisoni can be separated from O. martiensseni s.s. and O. madagascariensis based on greater inter-orbital width, braincase breadth and braincase height. The braincase of O. harrisoni is overall larger and higher than other Otomops spp. including O. martiensseni s.s.; this is most visible in the lateral view ( Fig. 10 View FIGURE 10 ). There is minimal overlap in braincase height (excl. tympanic bullae) between the corresponding sexes of O. harrisoni (males: 9.0– 9.8 mm; females: 8.7–9.5 mm) and O. martiensseni s.s. (males: 8.2–9.0 mm; females: 7.8–8.8 mm), and there is no overlap of O. harrisoni with O. madagascariensis (males: 7.9–8.5 mm; females: 7.9–8.2 mm). Compared to O. martiensseni s.s., the frontal region of the cranium of O. harrisoni is markedly more inflated giving it a dome-like appearance ( Fig. 10 View FIGURE 10 ). Conversely, O. martiensseni s.s. is characterised by a more anteriorly slanted frontal region, giving the braincase a lower lateral profile than O. harrisoni . The depression that lies across the fronto-parietal suture of the braincase is less pronounced in O. martiensseni s.s. than in O. harrisoni . The sagittal crest and lambdoidal crests are slightly more developed in O. harrisoni than in O. martiensseni s.s. and O. madagascariensis . They join above the occiputs to produce a “V-pattern”, giving the occipital region a rounded appearance; a characteristic feature of the genus. In O. harrisoni the sagittal crest remains slightly pronounced along the fronto-parietals, terminating at the interorbital depression by the junction of the nasals and frontals. In O. martiensseni s.s. and O. madagascariensis this feature is poorly developed. The anterior portion of the rostrum is more expansive in O. harrisoni than in O. martiensseni s.s. and O. madagascariensis . The mastoid projections are well-developed, elongate and contribute to the rounded appearance of the braincase when the cranium is viewed dorsally. In the lateral view, the foramen magnum of O. harrisoni is situated higher than in O. martiensseni s.s. The lacrimal process is distinct in most Otomops spp., but is particularly prominent in the Afro-Malagasy members including O. harrisoni . The jugal process of the zygomatic arch is thickened and the zygomaxillary junction is more laterally displaced than in O. martiensseni s.s. A prominent vertical process is located on the dorsal surface of the zygoma; it is slightly broader and rounder in appearance than in O. martiensseni s.s. and O. madagascariensis . The basisphenoid pits are elongate, oval-shaped, and deep. They appear to be wider in the holotype of O. martiensseni s.s. than in that of O. harrisoni . The broadened occipital condyles of the northeastern African species project outwards. The posterior border of the hard palate extends to the posterior margin of M3, similar to O. martiensseni s.s. Otomops harrisoni has external tympanic bullae that are longer and more posteriorly inflated than O. martiensseni s.s.
Typical of the genus, the mandible is thin and gracile, ending in an outwardly deflected angular process. The coronoid process is low-lying and the mandibular condyle is positioned in line with the lower toothrow. There is a distinct tubercle on the mandibular ramus between the third molar and the coronoid process. This tubercle has been observed in all three Afro-Malagasy species and has been reported from some Indo-Australasian species ( Kitchener et al. 1992). The dental formula of O. harrisoni is I 1 /2, C 1/1, P 2/2, M 3/3, characteristic of molossid bats. The upper incisors are moderately-developed and are separated from the canines by a slight diastema. This diastema is present in O. martiensseni s.s. and O. madagascariensis . The upper canines are long and fairly robust. The upper anterior premolar is small, reaching slightly past the cingulum of the posterior premolar, whereas the lower anterior premolar is half the size of the posterior premolar. The first two upper molars are similar in size, whilst the third molar is smaller in size and bears an unreduced commissure. The lower molars decrease in size from M1 to M3.
Molecular analyses. Analyses conducted using both mitochondrial and nuclear data are in agreement with regard to the number of extant clades/clusters within the Afro-Malagasy region. Genetic structure inferred by microsatellite analysis suggests strong genetic structure at regional (SECW, NEA), rather than population or colony level, and little gene-flow between the SECW and NEA regions, the habitats of O. martiensseni s.s. and O. harrisoni , respectively. The three groups formed, namely SECW, NEA and MAD ( Fig. 4 View FIGURE 4 ), correspond to the clades observed in the phylogram generated using concatenated cytochrome b and D-loop data ( Fig. 2 View FIGURE 2 ), the networks constructed using concatenated nuclear intron data ( Fig. 3 View FIGURE 3 ), as well as results reported by Lamb et al. (2008) and Richards et al. (2012). Clade G, comprising Malagasy Otomops specimens classified as O. madagascariensis , shows distinct and strongly supported (bootstrap support: 100%; posterior probability: 1.00) separation from the clades from mainland Africa ( Fig. 2 View FIGURE 2 ). Genetically, the sister lineages from mainland Africa and the Arabian Peninsula separate to form two independent reciprocally-monophyletic clades (subclades E and F), each having strong bootstrap and posterior probability support. These sister lineages are separated by a genetic p-distance of 2.10%. Nuclear sequence data supports this separation where, in figure 3, all Otomops spp. form independent networks at 95% confidence limit. The designation of O. martiensseni s.s. and O. harrisoni as separate species is supported based on the separation of individuals from the African mainland into 2 distinct groups derived from mitochondrial and nuclear sequence data and nuclear microsatellites.
Biology, distribution and conservation status. As mentioned above, specimens originating from Djibouti, Eritrea, Ethiopia, Kenya and Yemen have been assigned to this taxon. Therefore observations based on previously published literature ascribed to northeast African individuals now applies to O. harrisoni . While bioacoustic information is available from the literature for O. martiensseni s.s. ( Fenton & Bell 1981; Fenton et al. 2002, 2004; Adams et al. 2015) and O. madagascariensis ( Russ et al. 2003) , information on individuals attributable to O. harrisoni is uncertain. Limited time expansion recordings of a single hand-released individual captured at Bungule, Taita Hills in Kenya showed a frequency of maximum energy of 12.00 kHz, minimum frequency of 10.50 kHz, maximum frequency of 16.50 kHz, and a call duration of 9.00 ms ( Taylor et al. 2005); recorded echolocation parameters that lie within the previously described ranges of O. martiensseni s.s. ( Fenton & Bell 1981; Fenton et al. 2002, 2004; Adams et al. 2015) and O. madagascariensis ( Russ et al. 2003) . However, as Bungule is equidistant from the nearest sampled localities included within this study for both O. harrisoni (Chyulu Hills) and O. martiensseni s.s. (Usambara foothills), the taxonomic assignment of the Taita Hills individual is uncertain. To the best of our knowledge, no information on animals that can be definitively assigned to O. harrisoni is presently available.
Scat analysis of Ethiopian Otomops inferred that this is an insectivorous bat feeding predominantly (97% by volume) on medium (size range: 1–5 cm) to large (wing span size range: 2.5–30 cm) Lepidoptera of the moth families Noctuidae, Geometridae and Saturniidae ( Rydell & Yalden 1997; Heppner 2008). Otomops has a diet that chiefly comprises a single prey category and is thus highly specialized for moth predation through its jaw and tooth morphology, flight style and echolocation system ( Freeman 1981; Rydell & Yalden 1997; Jones & Rydell 2003). Otomops martiensseni s.s. bears similar morphological features as they relate to jaw architecture ( Harrison 1965; Monadjem et al. 2010). As with O. harrisoni , the diet of O. martiensseni s.s. is comprised mostly of Lepidoptera (MC Schoeman, pers. comm.). This is in sharp contrast to the diet of the insular O. madagascariensis that consists of varying proportions of Lepidoptera, Diptera and Coleoptera ( Andriafidison et al. 2007) .
The reproductive cycle of two Otomops populations in Kenya, corresponding to the known distribution of O. harrisoni , was investigated by Mutere (1973) at the sites of Suswa and Ithundu Caves. Results indicated that males reach sexual maturity around one year of age, weighing approximately 25 g, at which time a gular gland had developed. Sexual maturity in females is indicated by lactation and/or the presence of a foetus, at approximately one year of age and at a similar mass to males. The breeding season occurs once a year and pregnant bats were found from October through January, birthing mainly in December. Females have a gestation period of around 3 months and give birth to a single, hairless young. For more on the reproductive biology of Otomops individuals from Kenya, see Long (1995) and references therein.
Members of O. harrisoni are known from a variety of habitats in northeast Africa and the Arabian Peninsula, including woodlands and shrublands of the Arabian Peninsula and Eritrea; montane grasslands, woodlands and forests of Ethiopia; xeric grassland and shrublands of Djibouti; and the bushlands and thickets of Kenya ( Peel et al. 2007). According to the MaxEnt analysis conducted in this study ( Fig. 7 View FIGURE 7 A), this range may include additional localities within Uganda, northern areas of Tanzania and southern areas of Somalia. These bats are found at high altitudes (> 1000 m a.s.l) characterised by relatively drier climes (<500 mm annual precipitation), including warm semi-arid, tropical savanna, warm desert and, in the case of the Ethiopian highlands, temperate oceanic climates ( Peel et al. 2007). Given these preferences, individuals roost predominantly in mountain-associated cave systems and lava caves. Otomops is known to congregate in the lava caves of the Rift Valley at Mount Suswa as well as the Ithundu caves of the Chyulu Hills in Kenya ( Kingdon 1974; Kock et al. 2005) and is also found in the Sof Omar karst cave system of Ethiopia ( Largen et al. 1974), the Hud Sawa caves at the Al-Rayadi Al-Gharbi Mountains in Yemen ( Al-Jumaily 1999), the Day Forest National Park at the Goda Massif Mountains in Djibouti ( Hill & Morris 1971) and within a disused railway line tunnel near Asmara in Eritrea ( Kock & Zinner 2004). Otomops harrisoni roost sites are dark and poorly-ventilated, and consist primarily of natural structures with varying numbers of access points, e.g. whereas the tunnel housing the Asmara railway line has only one entry point ( Kock & Zinner 2004), the Sof Omar and Mount Suswa caves have over 30 entrances each ( Gunn 2004).
Otomops View in CoL are usually found roosting in large colonies, comprising smaller groups of several hundred, tightlypacked individuals. Colonies observed at the Sof Omar, Ithundu and well-studied Mount Suswa roosts have been estimated to contain up to 15 thousand individuals per site; however, it has been suggested that Mount Suswa colonies may exhibit migratory behaviour, periodically leaving the breeding site caves in which they are normally found ( Largen et al. 1974; Kock et al. 2005). At the time of publication, Al-Jumaily (1999) recorded the number of individuals in the Hud Sawa caves to be approximately 1500. Numbers in the Eritrean colony are comparatively lower than other roosts, i.e. approximately 500 individuals, since the railway tunnel would have only become suitable for habitation after 1974, making this a relatively young colony ( Kock & Zinner 2004).
The IUCN classification of currently-circumscribed O. martiensseni View in CoL is given as “Near Threatened” ( Mickleburgh et al. 2008) and possibly close to qualifying as “Threatened” as population numbers have decreased over time. In light of the recircumscription of O. martiensseni View in CoL , and the description of a new species, O. harrisoni View in CoL , the conservation of both of these species will have to be assessed in future studies. Ethiopia is signatory to a number of conventions including the Conservation of Migratory Species, which lists Otomops View in CoL as a species of interest. Although the Bale Mountains (Bale Mountains National Park and the Sof Omar National Monument included) are protected, protection measures are not clearly defined ( Vreugdenhil et al. 2012). The large colony at Mt Suswa in Kenya is subject to human disturbance within the caves and does not appear to be protected ( Kock et al. 2005). A protected area in the Day Forest National Park was assigned, although there may be a failure to implement prescribed conservation measures as a result of internal unrest in Djibouti ( Hutson et al. 2001; Magin 2001). Conservation initiatives for the protection of Otomops View in CoL within Yemen and Eritrea are unknown.
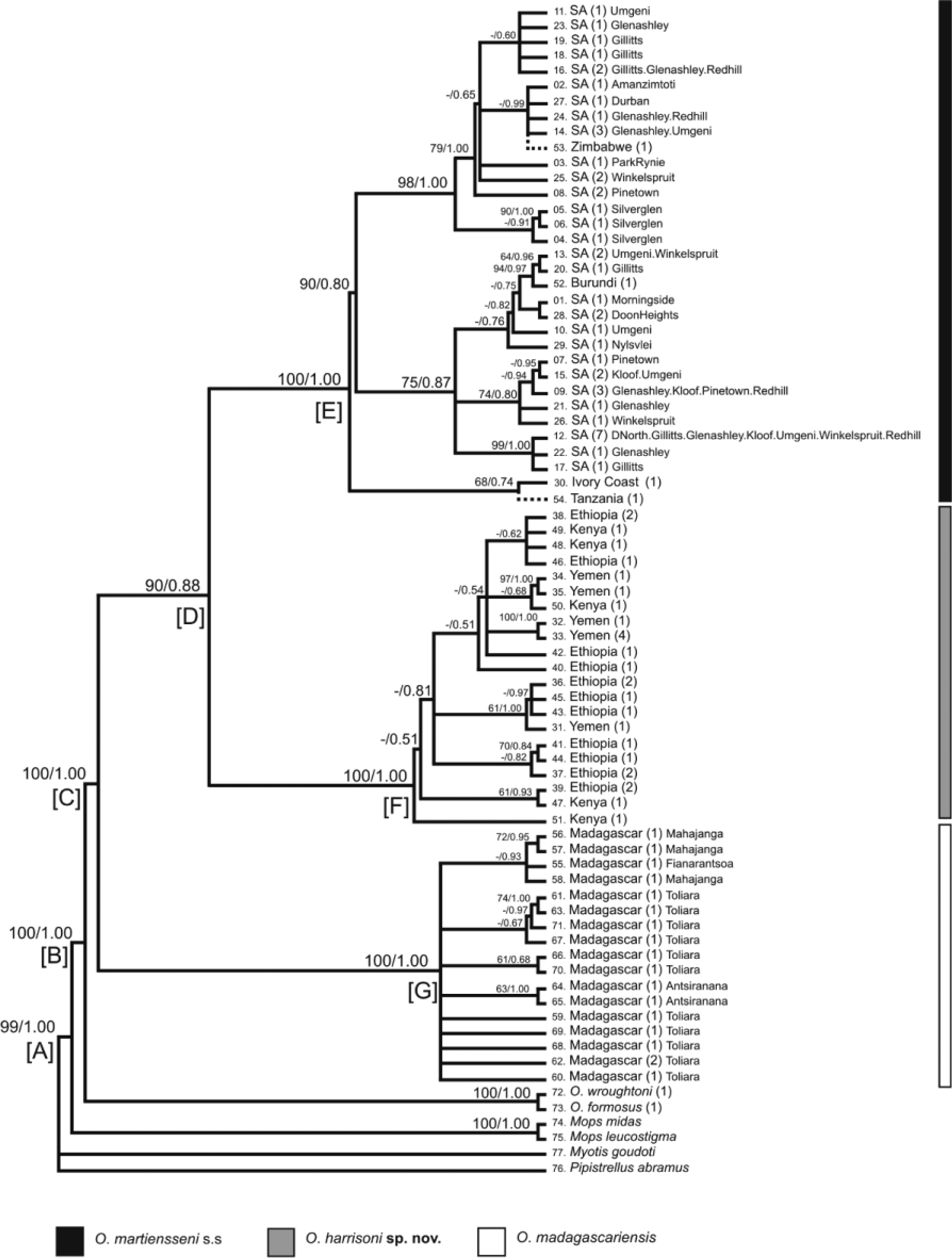
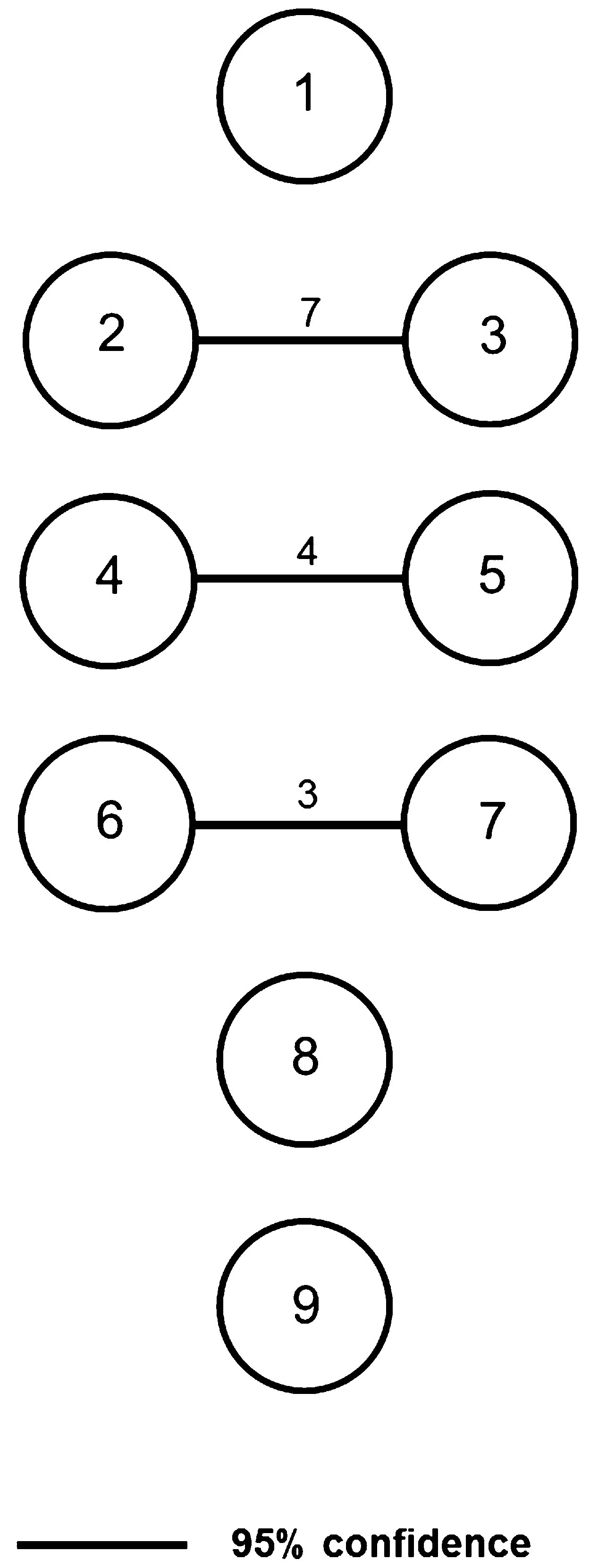
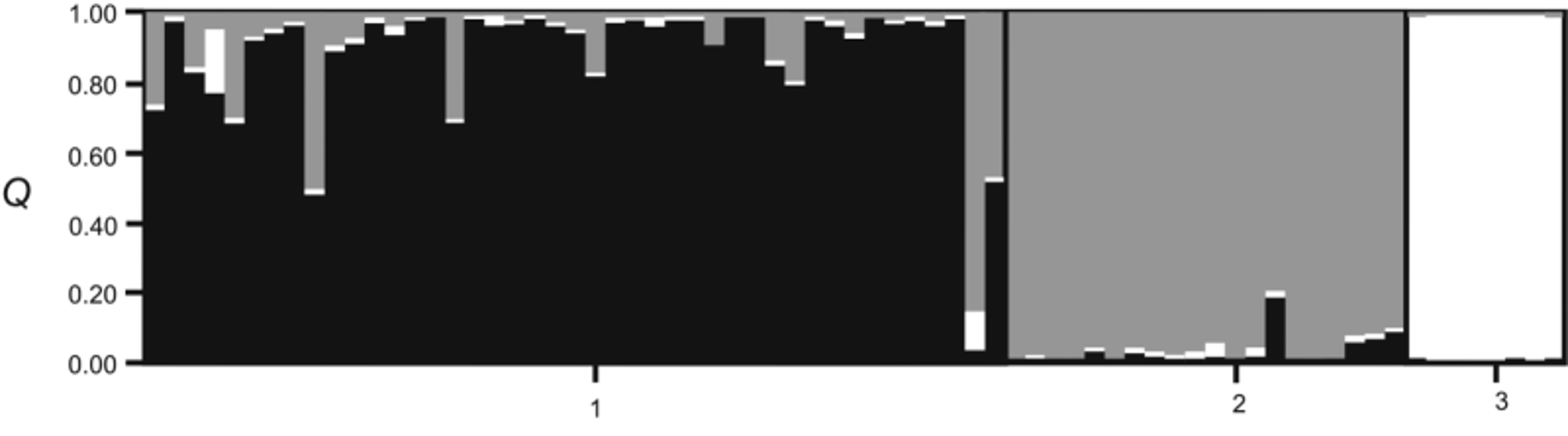
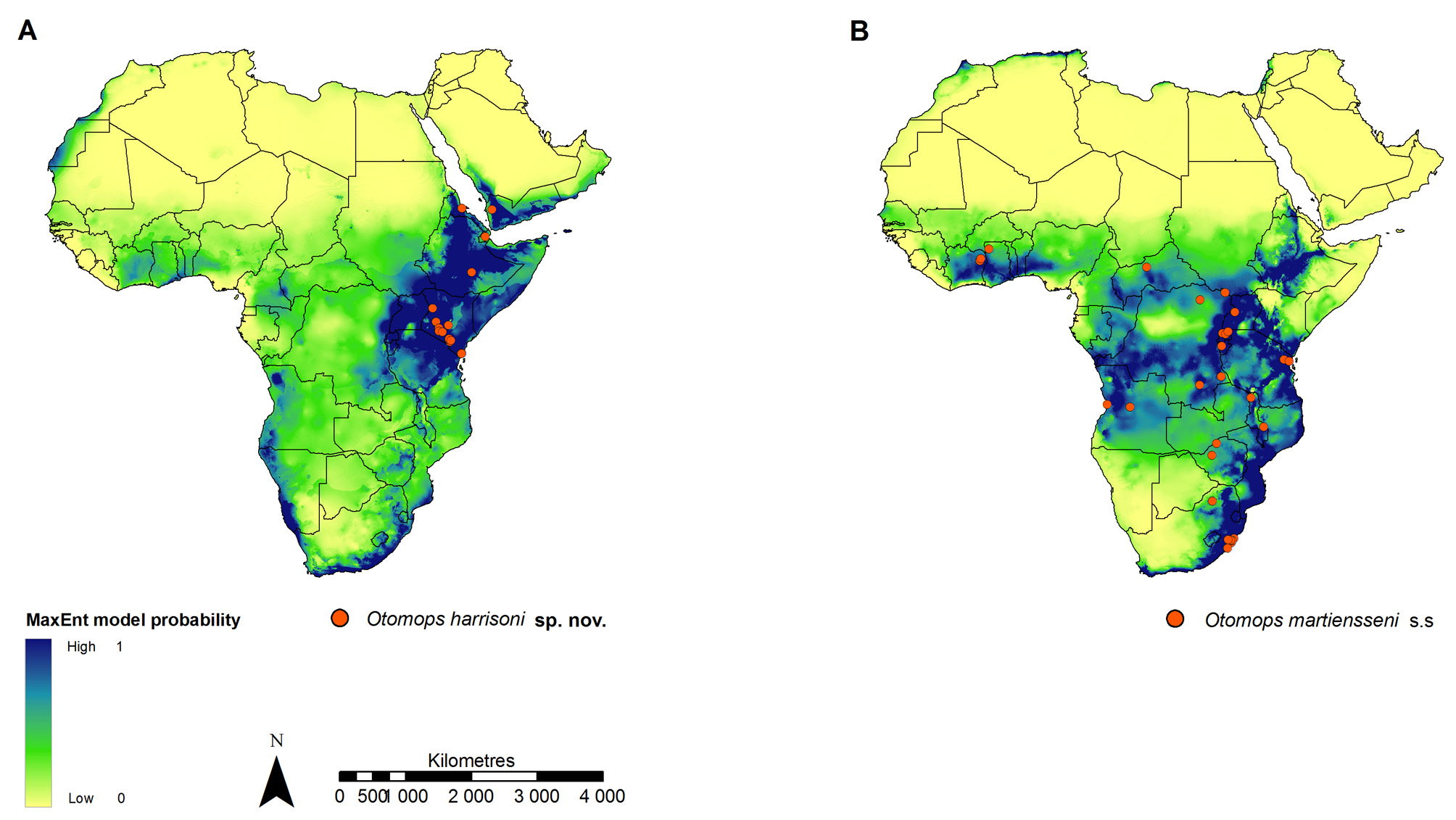
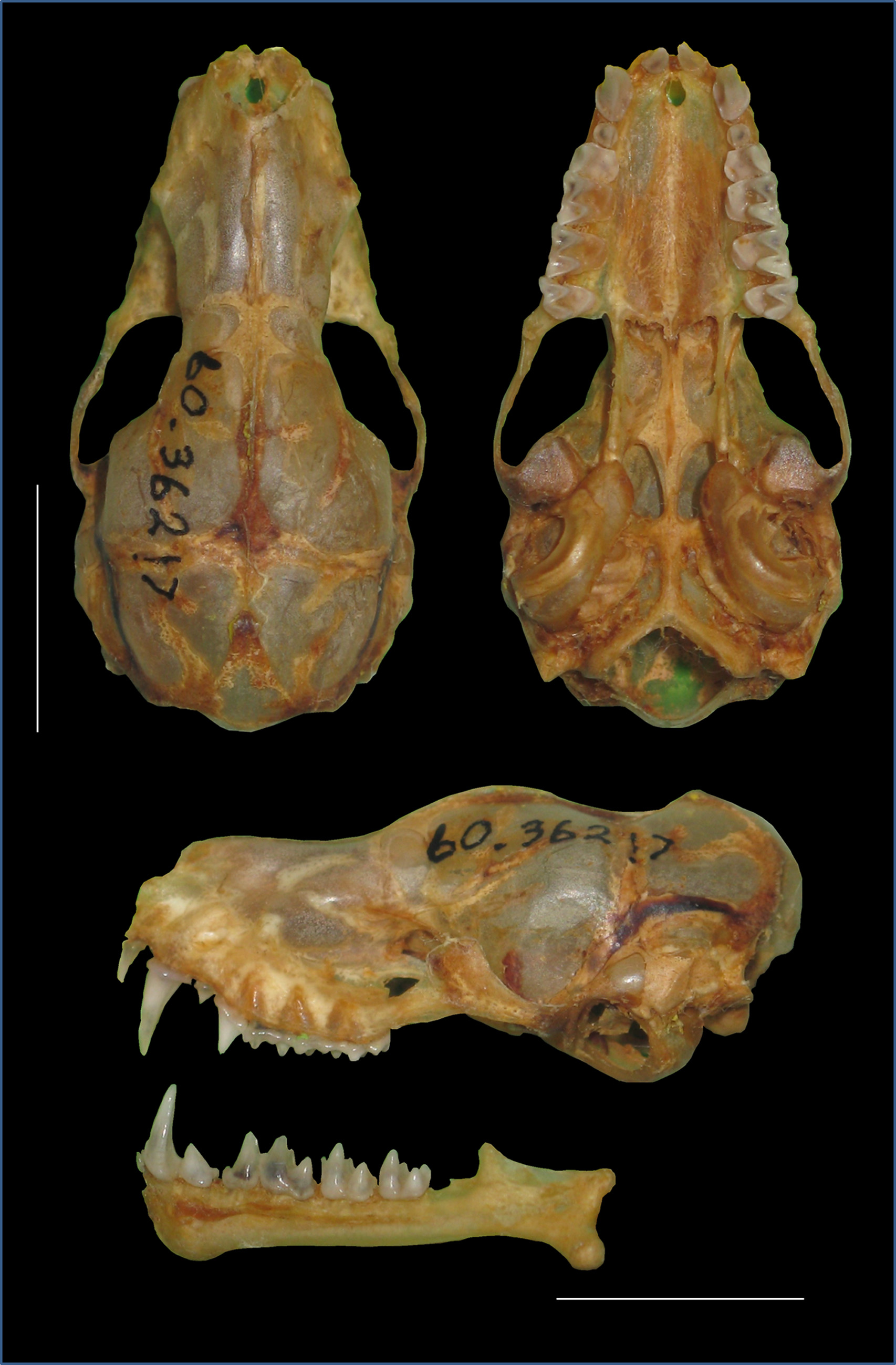
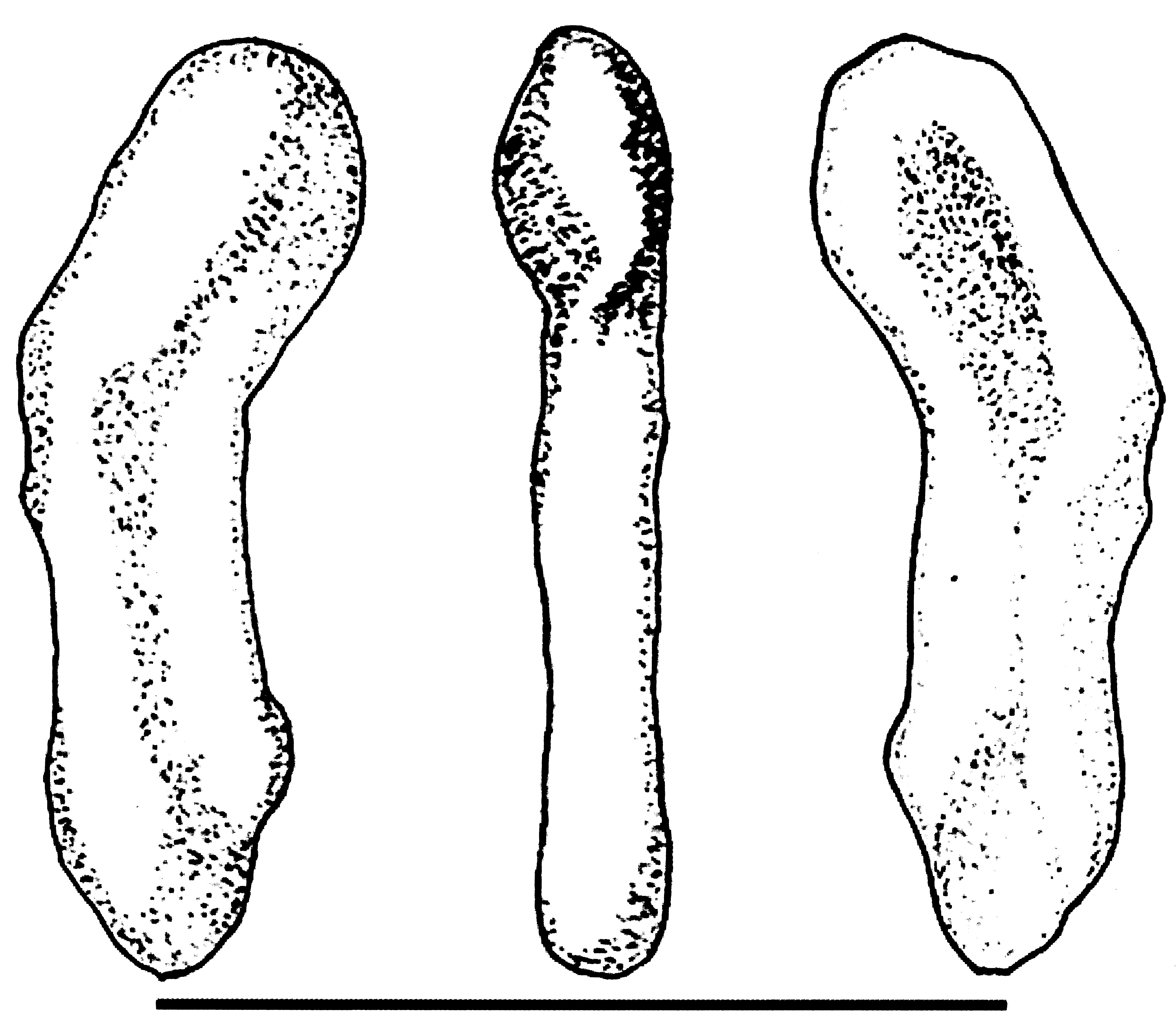

TABLE 4. Unique synapomorphies of O. harrisoni sp. nov. in the mitochondrial cytochrome b gene and D-loop region.
| Haplotype number 31 | Cytochrome b C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)C; T(831)C; C(837)T; T(933)C | D-loop A(4)T; A(6)G; C(41)T; C(58)T; A(59)C; A(101)G; - (131)A; T(140)C; A(142)G; G(144)A; T(167)C; A(177)G; T(182)C; C(200)T; T(202)C; T(205)G; C(220)T; A(235)G; G(250)A; C(252)T; T(264)C |
|---|---|---|
| 32 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)C; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; A(80)G; T(119)C; -(131)G; T(140)C; G(144)A; A(160)G; T(167)C; T(168)C; A(177)G; T(182)C; T(192)C; C(200)T; T(202)C; T(205)G; C(220)T; G(250)A; T(264)C |
| 33 | T (85) C; C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)G; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; A(80)G; T(119)C; -(131)G; T(140)C; G(144)A; A(160)G; T(167)C; T(168)C; A(177)G; T(182)C; T(192)C; C(200)T; T(202)C; T(205)G; C(220)T; G(250)A; T(264)C |
| 34 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)G; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; -(131)G; T(140)C; A(142)G; G(144)A; A(160)G; T(167)C; A(177)G; T(182)C; T(190)C; A(191)G; C(200)T; T(202)C; T(205)G; C(220)T; C(252)T; G(250)A; T(264)C |
| 35 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)G; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; -(131)G; T(140)C; A(142)G; G(144)A; A(160)G; T(167)C; A(177)G; T(182)C; T(190)C; C(200)T; T(202)C; T(205)G; C(220)T; G(250)A; C(252)T; T(264)C |
| 36 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(798)C; T(810)C; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; A(6)G; T(25)C; C(41)T; C(58)T; A(59)C; G(81)A; A(101)G; -(131)A; T(140)C; A(142)G; G(144)A; T(167)C; A(177)G; T(182)C; C(200)T; T(202)C; T(205)G; C(220)T; A(221)G; A(235)G; G(250)A; C(252)T; T(264)C |
| 37 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(798)C; T(810)C; T(831)C; C(837)T; T(933)C | A(4)T; T(5)C; A(15)G; C(41)T; A(59)T; -(131)G; T(140)C; A(142)G; G(144)A; T(166)C; G(176)A; A(177)G; T(182)C; A(195)G; C(200)T; T(202)C; T(205)A; A(206)G; C(220)T; G(250)A; T(264)C |
| 38 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)C; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; -(131)G; T(140)C; A(142)G; G(144)A; A(160)G; T(167)C; A(177)G; T(182)C; C(200)T; T(202)C; T(205)G; C(220)T; A(221)G; G(250)A; C(252)T; T(264)C |
| 39 40 | C(231)T; G(232)A; C(243)A; C(291)T; C(297)T; G(304)A; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(798)C; T(810)C; T(831)C; C(837)T; T(933)C C(231)T; G(232)A; C(243)A; A(258)C; C(291)T; C(297)T; C(336)G; A(426)G; C(564)T; A(651)G; T(664)C; A(786)G; T(810)C; T(831)C; C(837)T; T(933)C | T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; A(101)G; - (131)G; T(140)C; G(144)A; T(167)C; A(177)G; T(182)C; C(200)T; T(202)C; T(205)G; G(250)A; C(252)T T(2)C; A(4)T; C(41)T; C(58)T; A(59)T; A(101)G; - (131)G; T(140)C; A(142)G; G(144)A; A(160)G; T(167)C; A(177)G; T(182)C; C(200)T; T(202)C; T(205)G; C(220)T; G(250)A; C(252)T; T(264)C |
| HZM |
Museum of Natural History (Hrvatski Zooloski Muzej) |
| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| ROM |
Royal Ontario Museum |
| MRAC |
Musée Royal de l’Afrique Centrale |
| SMNS |
Staatliches Museum fuer Naturkund Stuttgart |
| NMP |
Natal Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Otomops harrisoni
| Ralph, Taryn M. C., Richards, Leigh R., Taylor, Peter J., Napier, Melanie C. & Lamb, Jennifer M. 2015 |
Otomops martiensseni: Kading et al., 2013 :2394
| Kading 2013: 2394 |
| Ralph 2013: 4234 |
| Tao 2013: 739 |
Otomops martiensseni:
| Kassahun 2015: 168 |
| Mortlock 2015: 1841 |
| Conrardy 2014: 259 |
| Yalden 2013: 480 |
Otomops martiensseni:
| Patterson 2012: 5 |
| Richards 2012: 913 |
Otomops martiensseni: Benda et al., 2011 :25
| Benda 2011: 25 |
Otomops martiensseni: Tong et al., 2009 :483
| Tong 2009: 483 |
Otomops martiensseni: Lamb et al., 2006 :46
| Lamb 2006: 46 |
Otomops martiensseni: Kock et al., 2005 :2
| Kock 2005: 2 |
Otomops martiensseni: Taylor et al., 2005 :26
| Taylor 2005: 26 |
Otomops martiensseni:
| Kock 2004: 3 |
| Debaeremaeker 2003: 221 |
| Jones 2003: 303 |
Otomops martiensseni: Pearch et al., 2001 :388
| Pearch 2001: 388 |
Otomops martiensseni:
| Al-Jumaily 1999: 241 |
| Rydell 1997: 72 |
Otomops martiensseni: Peterson et al., 1995 :178
| Yalden 1996: 91 |
| Peterson 1995: 178 |
Otomops martiensseni:
| Long 1995: 1 |
| Thollesson 1991: 26 |
Otomops martiensseni:
| Hickey 1987: 381 |
Otomops martiensseni martiensseni:
| Aggundey 1984: 144 |
| Fenton 1984: 398 |
| Freeman 1984: 400 |
Otomops martiensseni:
| Freeman 1981: 61 |
Otomops martiensseni:
| Norberg 1981: 365 |
Otomops martiensseni:
| Epelu-Opio 1974: 229 |
Otomops martiensseni martiensseni: Largen et al., 1974 :250
| Largen 1974: 250 |
| Kingdon 1974: 338 |
Otomops martiensseni: Warner et al., 1974 :171
| Valdivieso 1979: 6 |
| Kayanja 1978: 245 |
| Kayanja 1975: 166 |
| Warner 1974: 171 |
Otomops martiensseni martiensseni:
| Dulic 1973: 62 |
| Kinoti 1973: 129 |
| Hill 1971: 46 |
| Harrison 1965: 2 |