
Notohypsilophodon comodorensis Martínez, 1998
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3786.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:642CCCD6-95F7-4FFB-A7CE-10A3FF84F121 |
|
DOI |
https://doi.org/10.5281/zenodo.6137396 |
|
persistent identifier |
https://treatment.plazi.org/id/6B62B54B-5933-DA51-76D3-9A59232BFE05 |
|
treatment provided by |
Plazi |
|
scientific name |
Notohypsilophodon comodorensis Martínez, 1998 |
| status |
|
Notohypsilophodon comodorensis Martínez, 1998
Figs. 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10
Holotype. UNPSJB-Pv 942 ( Figs 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 ). A partial skeleton including four cervical vertebrae, seven dorsal vertebrae, four sacral vertebrae, six caudal vertebrae (all of the vertebrae are partially preserved), rib fragment, a portion of the left scapula, right coracoid, right humerus, left and right ulnae, an incomplete left femur, right tibia, an incomplete left tibia, left fibula, an incomplete right fibula; right astragalus, left calcaneum and 13 pedal phalanges.
Emended diagnosis. A small ornithopod dinosaur distinguished from all other basal ornithopods by the following autapomorphies originally proposed by Martinez (1998) and supported in this study: Anteromedial bulge on the proximal extremity of the tibia, calcaneum with a pronounced posterodistal projection. Additionally, the following new autapomorphies are proposed: Distal tip of the sacral neural spine strongly projected posteriorly, sternal process of the coracoids markedly reduced (see below).
Locality and horizon. 28 km northeast of the town of Buen Pasto, in the central-south of Chubut Province, Patagonia, Argentina ( Fig. 1 View FIGURE 1 ). Bajo Barreal Formation, Lower Member (Late Cretaceous, Cenomanian, see Martinez & Novas 2006).
Description and comparisons. As mentioned by Martínez (1998), the morphology of the material clearly differs from that seen in the most derived ornithopods (i.e., hadrosaurids). Likewise, the material does not possess synapomorphies or anatomical similarities of marginocephalians (i.e., pachycephalosaurs and ceratopsians). For this reason, members of both groups were excluded from our comparisons. Therefore, in order to ascertain the evolutionary affinities of Notohypsilophodon within Ornithischia , we compared the specimen to the following genera: basal neornithischians - Hexinlusaurus ( Barrett et al. 2005) and Yandusaurus ( He 1979; He & Cai 1984); basal (non-iguanodontian) ornithopods - Changchunsaurus ( Zan et al. 2005; Butler et al. 2011), Gasparinisaura ( Coria & Salgado 1996; Salgado et al. 1997), Gideonmantellia ( Ruiz-Omeñaca et al. 2012), Haya ( Makovicky et al. 2011), Hypsilophodon ( Galton 1974a; Torres & Viera 1994), Jeholosaurus ( Xu et al. 2000; Han et al. 2012), Koreanosaurus ( Huh et al. 2010) , Orodromeus ( Horner & Weishampel 1988) , Oryctodromeus ( Varricchio et al. 2007) , Parksosaurus ( Parks 1926; Galton 1973), Thescelosaurus ( Gilmore 1915; Galton 1974b; Boyd et al. 2009), Trinisaura ( Coria et al. 2013), and Yueosaurus ( Zheng et al. 2012); non-hadrosaurid iguanodontians - Anabisetia ( Coria & Calvo 2002) , Camptosaurus dispar ; Camptosaurus prestwichii = Cumnoria prestwichii , Camptosaurus aphanoecetes = Uteodon aphanoecetes [ Gilmore 1909; Galton and Powell 1980; Carpenter & Wilson 2008, see McDonald 2011), Dryosaurus ( Janensch 1955; Galton 1981), Iguanodon ( Norman 1980) , Macrogryphosaurus ( Calvo et al. 2007), Mantellisaurus ( Hooley 1925; Norman 1986; Paul 2006, 2008; McDonald 2012), Mochlodon ( Ösi et al. 2012), Ouranosaurus ( Taquet 1976) , Talenkauen ( Novas et al. 2004) , Tenontosaurus ( Forster 1990; Winkler et al. 1997), Valdosaurus ( Galton & Taquet 1982; Galton 2009; Barrett et al. 2011), and Zalmoxes ( Nopcsa 1900; Weishampel et al. 2003). Additionally, other specimens (e.g., the Australian ornithopods) as well as isolated and partially persevered materials are also used in the present comparison (see below).
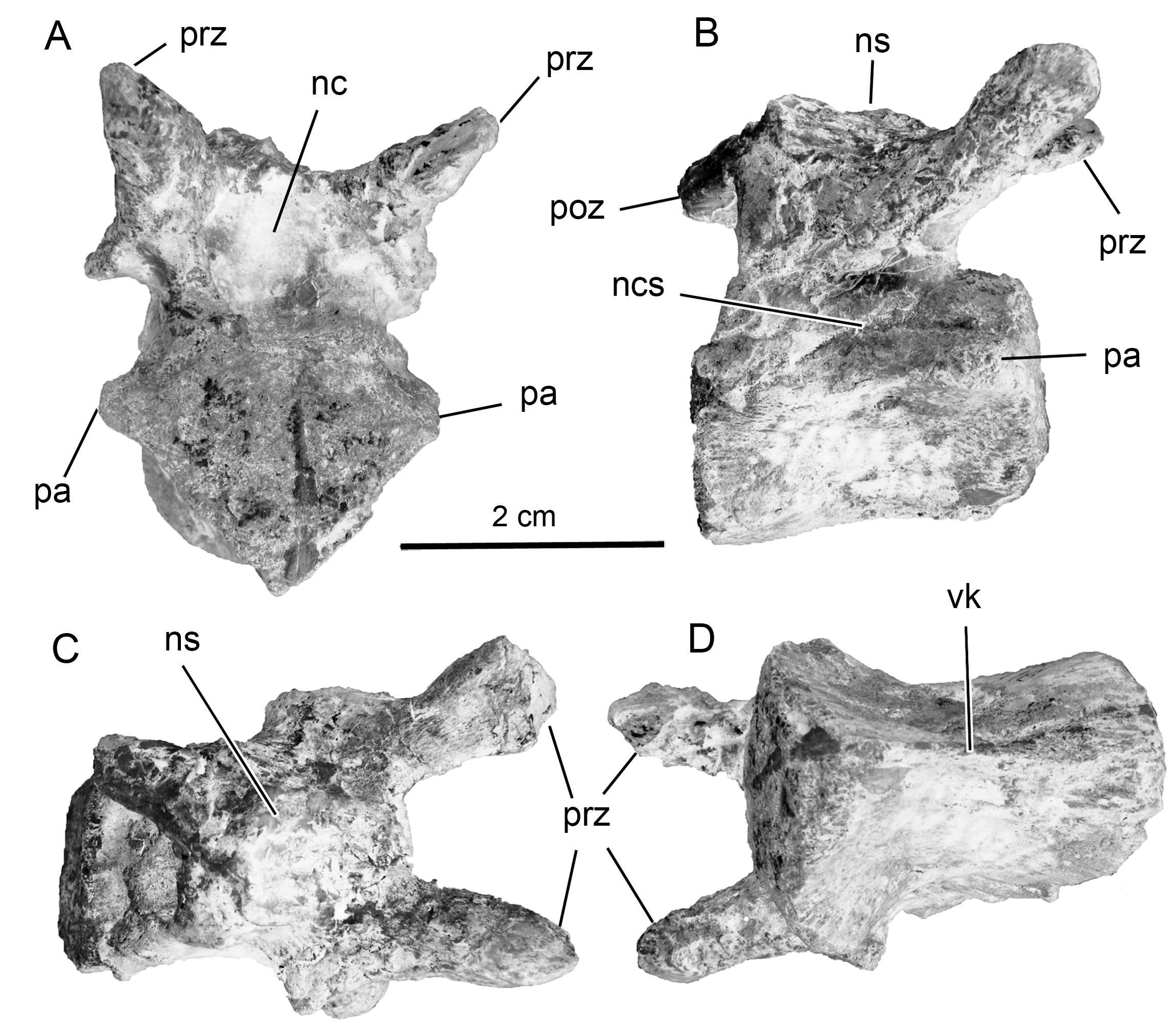
Cervical vertebrae. The cervical vertebrae ( Fig. 2 View FIGURE 2 ) of Notohypsilophodon comodorensis , based on the morphology of the centra and the portions of the neural arches preserved are presumably middle cervicals. The centra are amphyplatian as in Haya, Hypsilophodon and Jeholosaurus . In more derived ornithopods (e.g., Camptosaurus, Cumnoria , Dryosaurus , Tenontosaurus , Valdosaurus and Zalmoxes ), on the other hand, the cervical centra are slightly opisthocoelous or amphicoelous, as in Macrogryphosaurus. The centra of Notohypsilophodon are slightly wider than tall, as is the case in most basal neornithischians and basal ornithopods (e.g., Changchunsaurus , Hypsilophodon, Yueosaurus ). In ornithopods more derived than Camptosaurus (see Galton 2009: fig. 20), Macrogryphosaurus and Talenkauen , the cervical centra are anteroposteriorly longer than tall. In lateral view, the centra are “divided” in two slightly concave surfaces as in basal ornithopods due to the presence of the longitudinal ridge at approximately centrum midheight. Ventrally, the cervical vertebrae of Notohypsilophodon exhibit a thin midline keel similar to those seen in Anabisetia , Changchunsaurus , Gasparinisaura , Hypsilophodon, Macrogryphosaurus , and Valdosaurus (see Barrett et al. 2011: fig. 2) and unlike the thicker, rugose ventral keels, present in some basal iguanodontids (e.g., Camptosaurus, Cumnoria , Tenontosaurus, Uteodon , Zalmoxes: Carpenter & Wilson 2008 ; Norman 2004). In anterior view, the cervical vertebrae have pentagonal articular surfaces with an acute ventral apex as the rhabdodontid ornithopod Zalmoxes ( Weishampel et al. 2003: fig. 15) and the basal ornithopod Yueosaurus ( Zheng et al. 2012: fig. 3); however, in the latter, the ventral apex is more pronounced than in Notohypsilophodon and Zalmoxes . The neural canal is proportionally large and ovoid in outline (a feature which could be related to the juvenile status of Notohypsilophodon , see below). The articular facets of the prezygapophyses are subcircular in outline and project strongly anterodorsally as in Hypsilophodon . Based on the preserved portion of the cervical neural spines, they appear to be notably reduced, a condition also reported in the basal ornithopods Hypsilophodon , Gasparinisaura and the basal iguanodontian Talenkauen . In lateral view, the centra exhibit an open neurocentral suture, indicating that Notohypsilophodon was not somatically mature (as Martinez 1998, pointed out in his original study, see below). A parapophysis located just below the neurocentral suture is also present in Tenontosaurus and Thescelosaurus .
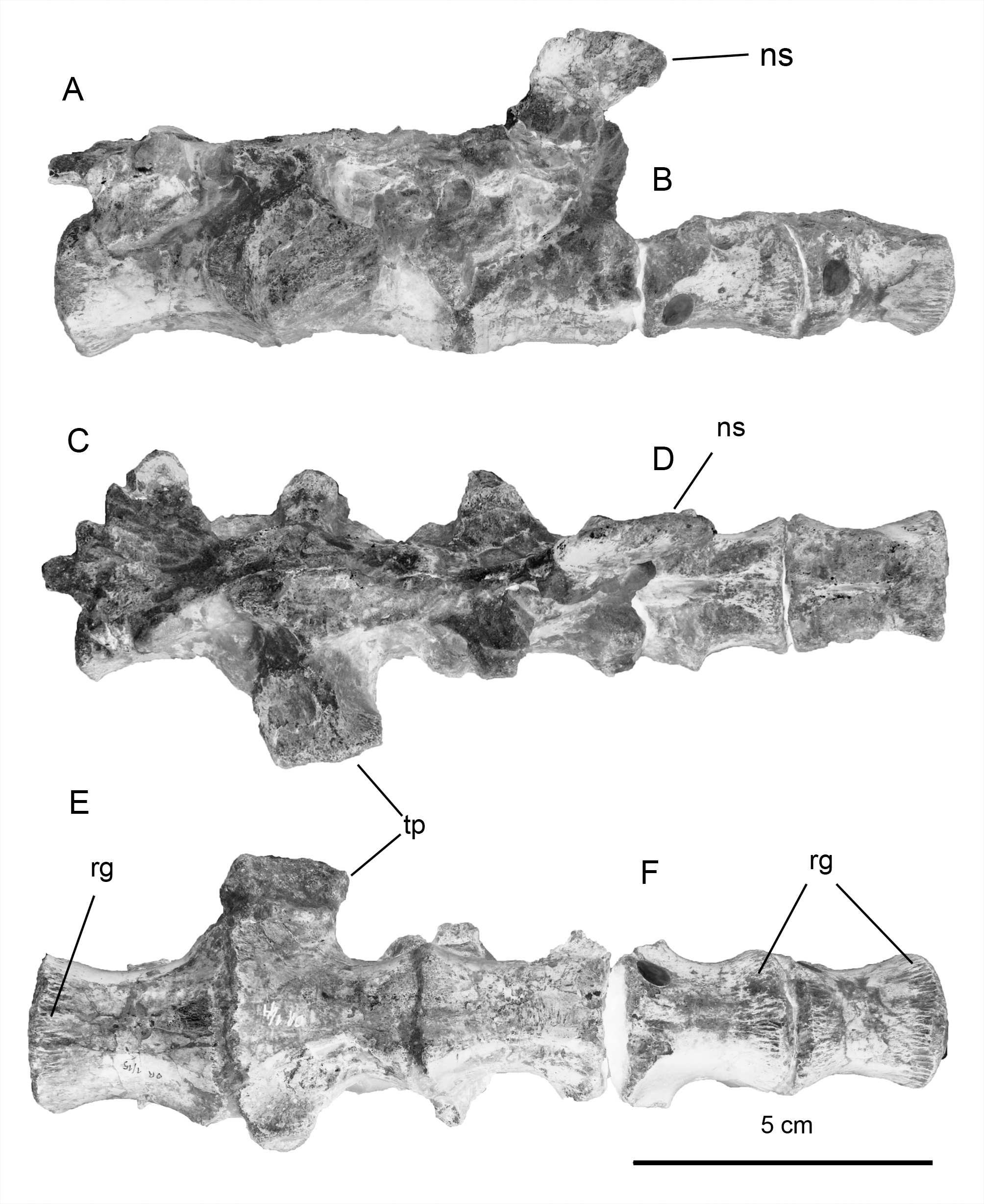
Dorsal vertebrae. The dorsal vertebrae ( Fig. 3 View FIGURE 3 ) are slightly amphicoelous and hourglass-shaped in ventral view as a consequence of their lateral compression. There are strongly-developed longitudinal striations around the margins of each of the articular surfaces, probably related to the insertion of soft tissues (i.e., the dorsal hypaxial musculature: Butler et al. 2010; Butler et al. 2011; Zheng et al. 2012). These longitudinal striations are comparable to those observed in many ornithopods, such as Camptosaurus , Dryosaurus, Macrogryphosaurus , Tenontosaurus , Thescelosaurus and a set of dorsal vertebrae recovered in Cretaceous outcrops of the Bajo Barreal Formation (Cenomanian-Turonian; UNPSJB-PV 960 see Ibiricu et al. 2010). Ventrally, the centra of Notohypsilophodon bear a midline keel as in the cervical vertebrae; however, this structure is not as sharp or as well defined as in the cervicals. A similar ventral keel is also seen in Changchunsaurus , Hypsilophodon, Trinisaura and Yueosaurus , whereas non-hadrosaurid iguanodontians (e.g., Cumnoria, Iguanodon , Mantellisaurus , Tenontosaurus , Zalmoxes and UNPSJB-PV 960) have centrum ventral surfaces that lack keels and which are more concave and rounded. The posterior surfaces of the centra are suboval in outline, a feature present in basal neornithischians (e.g., Hexinlusaurus and Yandusaurus ) and basal iguanodontians (e.g., Camptosaurus , Iguanodon and Mantellisaurus ). The transverse processes preserved in Notohypsilophodon project laterally, at approximately the same level as the zygapophyses, as is characteristic of basal ornithopods (Norman 2004; Makovicky et al. 2011). The preserved dorsal neural spine appears to be relatively low and anteroposteriorly narrow. This feature is comparable to the anatomy of basal ornithopods and differs from that seen in the more derived members of the group (i.e., nonhadrosaurid iguanodontians), where the dorsal neural spines are elevated and rectangular in shape (Norman et al. 2004).
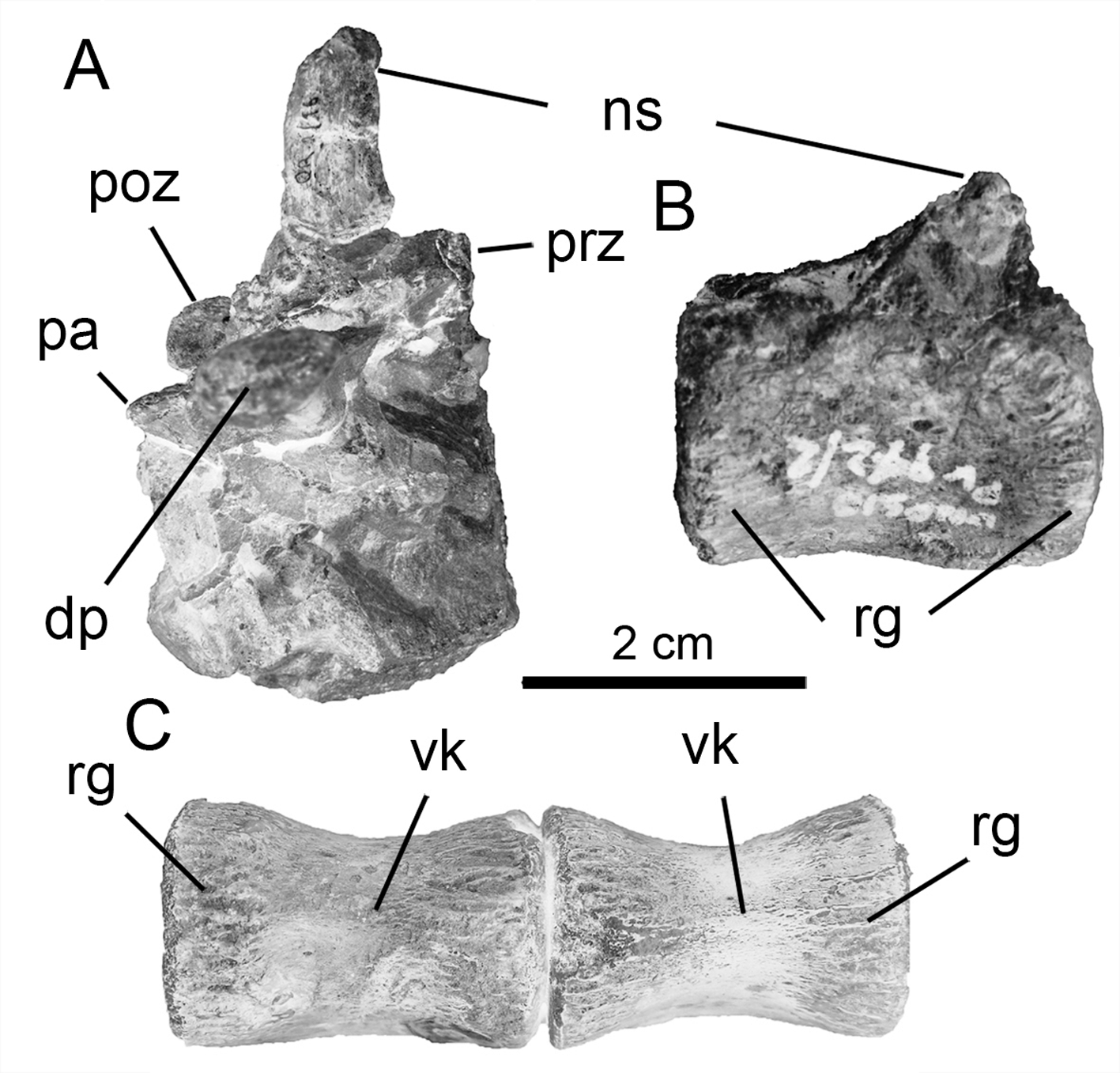
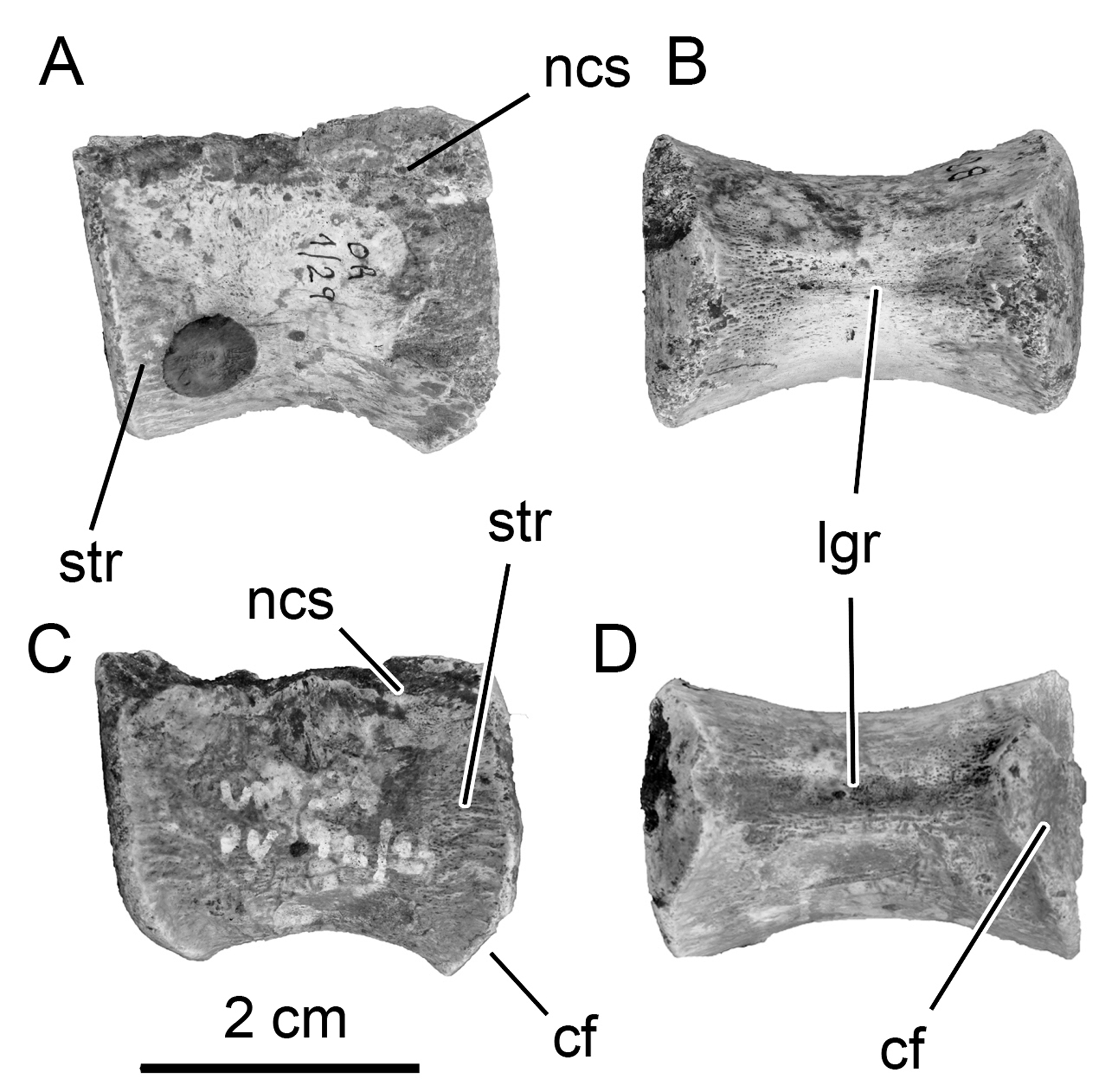
Sacral vertebrae. Due to the disarticulated nature of the sacral vertebrae, it is difficult to determine with confidence the positions of two of the sacral vertebrae recovered. Nevertheless, the sacral vertebra possessing a posterior articular surface that is laterally wider than its anterior counterpart is considered the dorsosacral ( Fig. 4 View FIGURE 4 , also see Butler et al. 2011). Unfortunately, it is not possible to determine the full sacral number (including the sacrodorsal vertebra) due to the absence of complete articulation. However, based on the preserved elements, we considered a putative number of five or six, as also known in many other ornithischians (e.g., Dryosaurus , Gasparinisaura, Haya, Hexinlusaurus , Hypsilophodon : also see Galton 1974a). On the other hand, with the exception of Oryctodromeus ( Varricchio et al., 2007) , some derived ornithopods, such as rhabdodontids (e.g., Zalmoxes and probably Mochlodon; see Ösi et al. 2012) and derived iguanodontians, possess more than six sacral vertebrae.
Of the two sets of sacral vertebrae recovered with Notohypsilophodon , three vertebrae, probably representing the anterior ones, are co-ossified ( Fig. 4 View FIGURE 4 A–C–E), whereas the other set, of two sacral vertebrae, are articulated but not co-ossified ( Fig. 4 View FIGURE 4 B–D–F). The sacral rib preserved (which is considered to be the first true sacral rib) is robust. The centra are slightly laterally compressed and are cotton-reel-shaped in ventral view. Circumscribing the articular surfaces, well-developed longitudinal striations are present. These striations continue onto the ventral surfaces of the centra, where they become more pronounced. The preserved sacral neural spine is wider anteroposteriorly than transversely. The distal end of the neural spine is posteriorly extended. In ventral view, the centra are comparable to those in Gasparinisaura , in possessing slightly concave ventral surfaces, whereas in many other basal ornithopods a longitudinal keel or ridge is present (e.g., Changchunsaurus and Haya).
Caudal vertebrae. The caudal vertebral centra ( Fig. 5 View FIGURE 5 ) are slightly amphicoelous, as in Gideonmantellia and UNPSJB-PV 960, and they are significantly longer anteroposteriorly than wide transversely. Additionally, comparisons with well-preserved caudal sequences (e.g., Hypsilophodon , Tenontosaurus ) demonstrate that these caudal vertebrae were located in the anterior portion of the tail. Notohypsilophodon possesses subrectangular intercentral articular surfaces, a feature shared with Tenontosaurus and UNPSJB-Pv 960. There are relatively welldeveloped longitudinal striations toward each lateral and ventral margin. This feature may be related to the insertion of soft tissue (e.g., the caudofemoralis musculature). Ventrally, the caudal centra of Notohypsilophon exhibit a marked longitudinal groove and well-developed facets for the insertion of the haemal arches. These last two features are also seen in several basal ornithopods and basal non-hadrosaurid iguanodontians (e.g., Gasparinisaura , Hypsilophodon, Macrogryphosaurus , Zalmoxes , UNPSJB-Pv 960 and, although less developed, in the basal ornithopod Jeholosaurus ). The presence of the marked longitudinal groove could be related to either ontogenetic status ( Han et al. 2012) or sexual dimorphism ( Nopcsa 1929; Ibiricu et al. 2010). No ossified tendons are preserved in Notohypsilophodon , which may be related to preservation rather than genuine absence.
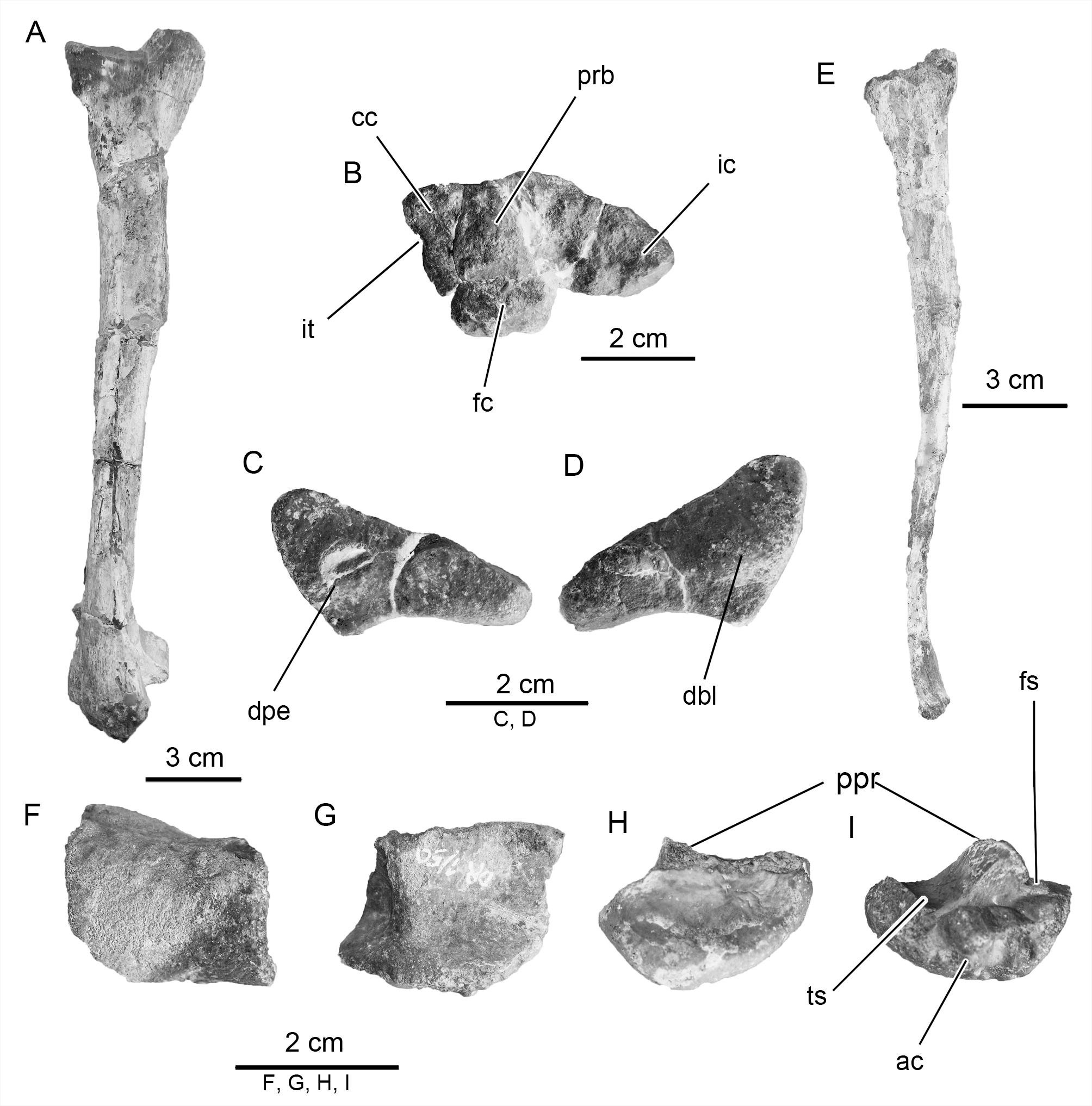
Scapula. The left scapula of Notohypsilophodon ( Fig. 6 View FIGURE 6 A–B; Table 1) preserves the proximal end and a portion of shaft, which is transversally narrow as in Gasparinisaura . The scapula and coracoids are unfused as in the majority of small ornithischians ( Han et al. 2012), and unlike Koreanosaurus and Oryctodromeus . The proximal end is expanded as in basal ornithopods (e.g., Gasparinisaura , Hypsilophodon ) and basal iguanodontians (e.g., Dryosaurus , Tenontosaurus ). The acromial process is not complete; therefore, it cannot be determined if it process was well developed, as in Anabisetia , or only moderately developed, as in Gasparinisaura ( Coria & Calvo 2002) . In dorsal view, the scapula of Notohypsilophodon exhibits two asymmetrical surfaces, the glenoid fossa and the coracoid articulation. The former is concave, whereas the latter is gently convex. The glenoid fossa is subrectangular in outline. Likewise, dorsally to this fossa, a marked concavity (= supraglenoid fossa; Zheng et al. 2012) is also present. This last feature is shared with various ornithischians including: Hexinlusaurus, Hypsilophodon , Lesothosaurus ( Thulborn 1972) , Thescelosaurus , Tenontosaurus, Uteodon , and Yueosaurus. However, the supraglenoid fossa in Notohypsilophodon is more pronounced than in all of these taxa with the exception of Tenontosaurus , whose fossa is similar to that seen in Notohypsilophodon ( Martinez 1998) .
Element Measurement Humerus Total length 127.3 Ulna Total length 91.3 Femur (proximal end) Maximun anteroposterior width 44.9
Maximun lateral width 47.7 Femur (distal end) Maximun anteroposterior width 45.5 Tibia Total length 211.5
Greatest proximal width 56.5
Greatest distal width 45.5 Fibula Total length 192
Greatest proximal width 29.6 Greatest distal width 14.1 Phalanx (?-1) Total length 36.1 Coracoids. The semicircular shape of the right coracoid ( Fig. 6 View FIGURE 6 C), with a slightly convex lateral surface and medially concave surface, is comparable to that of Gasparinisaura , Hypsilophodon and Yandusaurus . Conversely, in Changchunsaurus , Jeholosaurus and Koreanosaurus , the coracoids are sub-rectangular in shape with flat surfaces. In Trinisaura, the coracoid is semicircular in outline, as in Notohypsilophodon ; however, in the former the dorsal margin is interrupted by a marked concavity (see Coria et al. 2013: fig. 3). The sternal process, in Notohypsilophodon is not as well-developed as those of Anabisetia , Gasparinisaura, Haya , Koreanosaurus , Tenontosaurus or Trinisaura. Although, it is taphonomically affected on its lateral size, in Notohypsilophodon the sternal process is markedly more reduced than in those ornithopods (see below).
The coracoid foramen is circular in outline, proportionately large and completely enclosed. The coracoid foramen in Notohypsilophodon is placed well above the scapula-coracoid suture (i.e., there is no contact between the foramen and scapula). The shape and position of the coracoid foramen is similar to those of Anabisetia , Gasparinisaura, Trinisaura and Tenontosaurus (see Winkler et al. 1997: fig. 16a), and unlike that observed in Hypsilophodon . In the latter taxon, the foramen is positioned close to the suture and connected to the scapula by a groove ( Galton 1974a; Huh et al. 2010). In more basal forms (e.g., Changchunsaurus , Koreanosaurus ), the foramen is smaller than in Notohypsilophodon and located more towards the center of the coracoid plate.
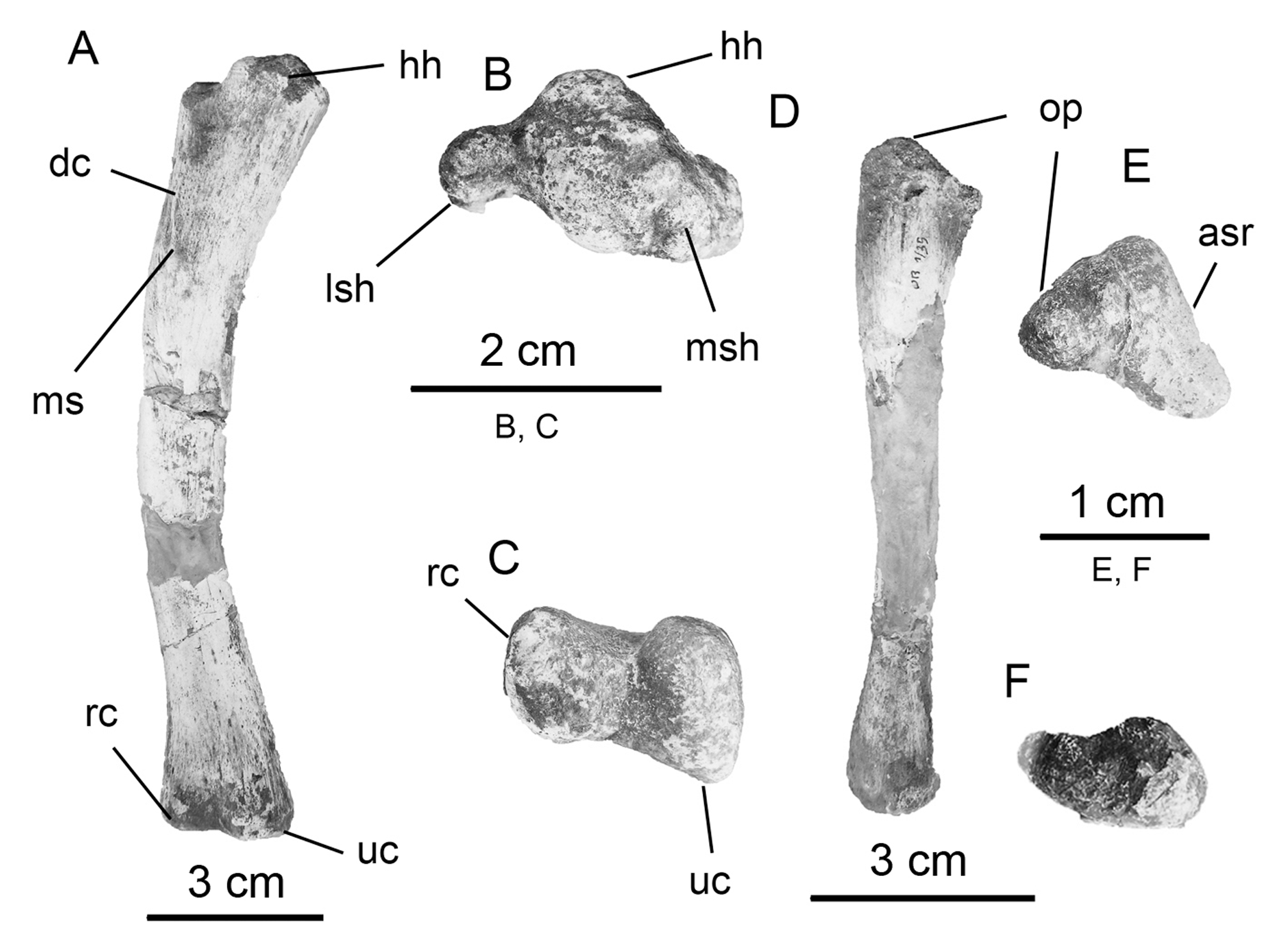
Humerus. The right humerus ( Fig. 7 View FIGURE 7 A–C; Table 1) of Notohypsilophodon is gracile (1.5 cm, measurement of the shaft taken in anterior view below the deltopectoral crest) as in Anabisetia , Gasparinisaura , Talenkauen and Trinisaura, whereas in the rhabdodontid Zalmoxes (approximately 3 cm, measurement taken from Weishampel et al. 2003: fig. 20d) and the basal iguanodontian Tenontosaurus , the humerus is robust (Norman et al. 2004). The shaft is slightly twisted, as is characteristic of a variety of basal ornithopods (Norman et al. 2004). Nevertheless, the rotation of the humeral shaft appears to be less than in Hypsilophodon ( Galton 1974a: figs. 38–39) and resembles the condition of Anabisetia . In anterior view, the shaft is bowed as in Yandusaurus and unlike those of Anabisetia and Trinisaura where the bowing is less marked. The deltopectoral crest is reduced (i.e., to shallow rugosities for the pectoral muscle insertions) and is positioned in the proximal portion of the humerus as in Anabisetia , Bugenasaura (Norman et al. 2004) , Gasparinisaura , Talenkauen and Trinisaura. By contrast, the deltopectoral crests of basal neornithischians and ornithopods (e.g., Haya, Hypsilophodon , Jeholosaurus , Koreanosaurus , Yandusaurus , among others) are well developed and angular in outline. Likewise, the deltopectoral crest in basal (non-hadrosaurid) iguanodontians is robust and it occupies approximately 50% of total humerus length (Norman et al. 2004). The proximal end of the humeral head is transversally expanded as in Anabisetia and Gasparinisaura , and exhibits a marked tuberosity, similar to that described in Dryosaurus ( Martínez 1998) . Thus, the proximal end is wider than the distal end as in Anabisetia and Trinisaura. Conversely, in Hypsilophodon , Yandusaurus and basal iguanodontians (e.g., Tenontosaurus ), the proximal and distal ends of the humerus are approximately equal in mediolateral width. The distal end of the humerus exhibits moderately developed radial and ulnar condyles, as well as an intercondylar groove (=fossa) on its anterior surface. This condition differs from that seen in Hypsilophodon , Thescelosaurus and non-hadrosaurid iguanodontians (e.g., Camptosaurus , Iguanodon , Mantellisaurus ) where the condyles and the groove are more strongly developed. The lateral radial condyle is expanded anteroposteriorly and the medial ulnar condyle is rounded, both of which are features that Notohypsilophodon shares with basal neornithischians and ornithopods such as Yandusaurus and Koreanosaurus .
Ulna. The ulna ( Fig. 7 View FIGURE 7 D–E–F; Table 1) of Notohypsilophodon is slender, as in Hypsilophodon , in contrast to the more robust ulnae of non-hadrosaurid iguanodontians. In proximal view, the ulna is triangular in outline (formed by the head of the ulna and the lateral and medial processes according to Butler et al. 2011) with the lateral border convex, the medial slightly concave and the articular surface for the radius gently concave. The olecranon process is reduced, and comparable to that observed in Hypsilophodon and Parksosaurus . Anabisetia , Dryosaurus and a variety of basal ornithopods ( Huh et al. 2010; Makovicky et al. 2011) possess relatively prominent olecranon processes that are more pronounced than that present in Notohypsilophodon , but reduced in comparison with those of heterodontosaurids (Norman et al. 2004). The shaft of the ulna is narrow toward the medial section and slightly expanded distally. The ulna is approximately 75% of the length of the humerus. These last two features in Notohypsilophodon are shared with basal ornithopods.
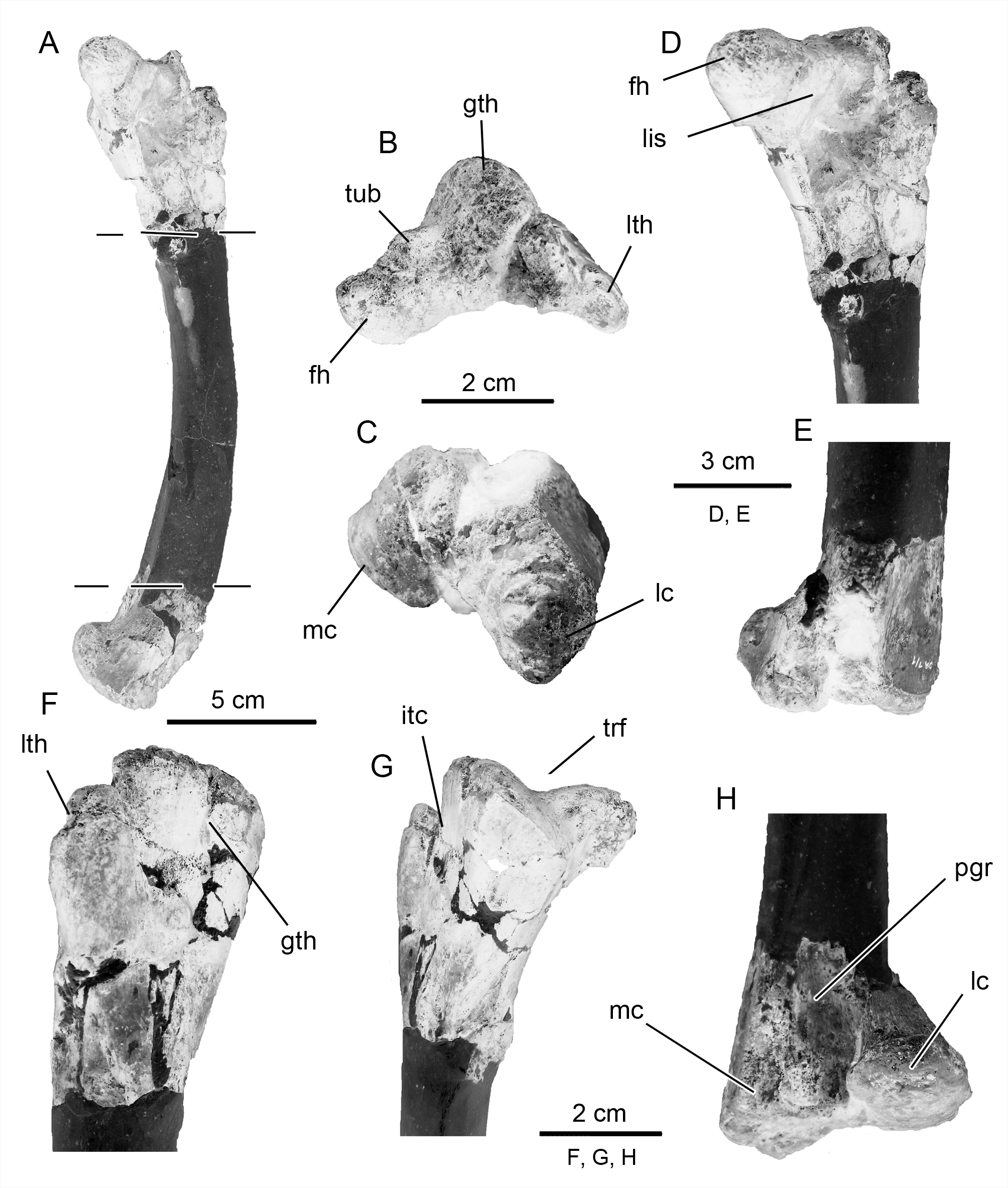
Femur. The proximal and distal ends of the left femur ( Fig. 8 View FIGURE 8 , Table 1), and a portion of the shaft, have been recovered in Notohypsilophodon . The preserved portion of the shaft strongly suggests that it was slender (a constricted neck is present) as in Gasparinisaura and other basal ornithopods such as Hypsilophodon , Jeholosaurus , Koreanosaurus and Yueosaurus. The femoral head is bulbous (i.e., with a well-developed spherical articular condyle), a feature shared with Gasparinisaura . In Notohypsilophodon , as in Orodromeus , the femoral head is oriented slightly dorsally with respect to the shaft, forming an angle greater than 100°, unlike the angle of approximately 90° found in most basal ornithopods (Norman et al. 2004). On the other hand, overall, the femora of basal iguanodontians (see Naish & Martill 2001: figs. 1-3), including rhabdodontids and ankylopollexians, are more robust than those of Notohypsilophodon , with the exception of those in dryosaurids, which are slender (e.g., Callovosaurus [see Ruiz-Omeñaca et al. 2007], Dryosaurus and Valdosaurus [see Barrett et al. 2011]). In proximal view, the head is compressed anteroposteriorly and the trochanters are expanded anteroposteriorly, as in Changchunsaurus . The femoral head is separated from the trochanters by a shallow depression, the fossa trochanteris. This fossa is absent in basal ornithischians such as Hexinlusaurus and heterodontosaurids, whereas it is usually present in cerapodans (Butler et al. 2011). Likewise, in proximal view, a well-marked convexity (i.e., “tuberosity” of Novas 1996) and a shallow notch (=sulcus, sensu Butler et al. 2011) for the ligamentum capitis femoris are present in Notohypsilophodon . The greater trochanter is anteroposteriorly expanded and its dorsal surface is slightly convex, whereas the “finger-like” lesser (=anterior) trochanter is also gently convex in the same direction. The greater and lesser trochanters are unfused and separated by a marked cleft (the intertrochanteric cleft for some authors, for the m. iliofemoralis insertion: Hutchinson & Gatesy 2000), a feature that occurs in Gideonmantellia, Hexinlusaurus, Hypsilophodon , Valdosaurus and Yandusaurus (Norman et al. 2004; Barrett et al. 2011; Ruiz- Omeñaca et al. 2012). Additionally, this feature is variable in Hypsilophodon (P. M. Barrett, pers. comm., December 2013). In contrast, in Gasparinisaura , Koreanosaurus , and Yueosaurus, the greater and lesser trochanters are fused. Although in Anabisetia, Trinisaura (see Coria et al. 2013), a cleft separating those trochanters is present, it is narrow in comparison with that seen in Notohypsilophodon . In lateral view, the tip of the lesser trochanter is set below the level of the greater trochanter, as in most basal ornithischians ( Agnolin et al. 2010). This differs, for example, from those see in Anabisetia , Changchunsaurus , Jeholosaurus , Koreanosaurus and dryosaurids (both trochanters are nearly to the same level, also see Naish & Martill 2001: fig. 1-6, plate 8). The distal end of the femur in Notohypsilophodon exhibits two asymmetrical condyles, the tibial (medial) and fibular (lateral). The former is larger than the latter and the rounded articular surfaces are covered with well-developed rugosities for soft tissue insertions. The lateral condyle projects slightly laterally, like those of Gasparinisaura and Trinisaura. Lateral to the medial condyle, a marked and slightly concave surface is present, as in Hypsilophodon (i.e., the extension of the medial surface occupies all the lateral aspect of the tibial condyle). Notohypsilophodon possesses a shallow anterior (extensor) groove as in basal euornithopods (Norman et al. 2004). This groove is absent in some other taxa (e.g., Gasparinisaura ) but well-developed in Anabisetia , Muttaburrasaurus ( Bartholomai & Molnar 1981; Molnar 1996 also see Agnolin et al. 2010) and Trinisaura. The posterior (flexor) intercondylar groove is comparable to those observed in basal ornithopods (Norman et al. 2004), whereas in basal non-hadrosaurid iguanodontians it is wider. Lastly, the distal end of the “Victoria femur type I” ( Rich & Rich 1999) is similar in morphology to that of Notohypsilophodon , in having a laterally placed fibular condyle, transverse compression and a medial condyle that is markedly larger than the lateral ( Agnolin et al. 2010). However, these features probably are due to taphonomic aspects rather than genuine anatomical characteristics (P. M. Barrett, pers. comm., December 2013).
Tibia. The straight right tibia ( Fig. 9 View FIGURE 9 A–B–C–D; Table 1) of Notohypsilophodon is slender, a feature shared with a variety of ornithopods (e.g., Anabisetia , Gasparinisaura, Gideonmantellia , Hypsilophodon, Yueosaurus , among others). The proximal end is anteroposteriorly expanded as in Changchunsaurus and Gasparinisaura . The cnemial crest is modestly developed and slightly laterally and dorsally projected as is usual in basal ornithopods ( Huh et al. 2010). The cnemial crest is separated from the fibular condyle by a groove (the incisura tibialis see Butler et al. 2011) as in Koreanosaurus , Jeholosaurus and Hypsilophodon . Both condyles of Notohypsilophodon , the rounded inner and fibular are anteroposteriorly expanded, particularly the latter. The proximal expansion of the condyles is more strongly developed than in basal forms such as Haya and Koreanosaurus . Proximally, the tibial surface exhibits a well-developed bulge. The distal end of the tibia in Notohypsilophodon , is subtriangular in outline as in Dryosaurus and Yueosaurus. Additionally, it is transversely expanded as in basal ornithischians ( Makovicky et al. 2011). The outer malleolus is larger than the inner malleolus, a condition reported in basal ornithopods ( Zheng et al. 2012) and unlike the subequally-sized malleoli in Gasparinisaura . With regard to the distal end, the subtriangular shape, prominently extended anteromedial flange, presence of a shallow lateral groove, and the similarly-shaped astragalar articular surface, are features that Notohypsilophodon shares with Dryosaurus altus (see Novas 1996: fig. 4I). Also in distal view, the articular surface of the tibia exhibits a welldeveloped perforation similar to that drawn in Dryosaurus (see Novas 1996: fig. 4I). This sub-oval distal perforation, which could be related with insertion of soft tissue (probably variable ossification of the cartilage capping the tibia), is placed closer to the external margin of the inner malleolus than the outer malleolus. Likewise, its counterpart, in the same position, exhibits a well-developed distal bulge ( Fig. 9 View FIGURE 9 C and D).
Fibula. The left fibula ( Fig. 9 View FIGURE 9 E) of Notohypsilophodon is slender. Its proximal end is anteroposteriorly expanded and its surface is slightly concave. The shaft narrows somewhat to the mid-length of the bone. At this point, the shaft exhibits a thin well-developed flange (flange on medial surface of fibula according to Butler et al. 2011) that extends medially. This flange reaches the distal end of the fibula. The distal end is moderately expanded.
Astragalus. The partially preserved right astragalus ( Fig. 9 View FIGURE 9 F–G) has a sub-rectangular shape. In distal view, it is convex, whereas in dorsal view, it is concave and divided into two articular surfaces divided by a ridge. The surface that received the distal end of tibia is wide, whereas the laterally placed surface for contact with the calcaneum is small. The ascending process is sub-triangular in shape and it is moderately developed, as also described in other basal ornithopods (Norman et al. 2004).
Calcaneum. The left calcaneum ( Fig. 9 View FIGURE 9 H–I) is semicircular in shape, slightly concave laterally, and mediolaterally compressed. The articular surfaces for contact with the distal ends of tibia and fibula are divided by a moderately developed and partially oblique proximal projection. The fibular and tibial articular surfaces are concave. The tibial articular surface is open medially. In contrast, the fibular surface is delimited by a thin lip. The medial surface of the calcaneum displays a well-developed astragalar process or contact, as well as a medial groove. Distally, this element exhibits well-developed rugosities indicating soft tissue insertions. The semicircular shape is shared with other ornithopods including Camptosaurus, Cumnoria ; Gasparinisaura , Hypsilophodon , Iguanodon , Ouranosaurus , Valdosaurus , Zalmoxes , and UNPSJB-PV 960, among others. Likewise, the proximal ridge that separates the fibular and tibial articular surfaces, the well-developed astragalar process, and the medial groove are also anatomical characteristics shared with these other ornithopods. However, by contrast, the medial surface of the calcaneum in Changchunsaurus is flat, lacking the groove and process. The tibial articular surface in Notohypsilophodon is concave in lateral view and it may have distally enveloped the anterior portion of the tibial lateral malleolus, this condition, when in natural articulation, is comparable to those seen in Gasparinisaura , Hypsilophodon , Ouranosaurus and UNPSJB-PV 960. In Notohypsilophodon there is a marked difference in size between the fibular and tibial articular surfaces, the former being smaller than the latter in lateral view. This feature is shared with Gasparinisaura and UNPSJB-PV 960, but in Camptosaurus, Hexinlusaurus , Hypsilophodon and Zalmoxes these fossae are subequal in size in the same view. Likewise, in iguanodontians such as Iguanodon and Mantellisaurus , the fibular articular surface is clearly longer than the tibial (see Ibiricu et al. 2010), unlike the condition in Notohypsilophodon .
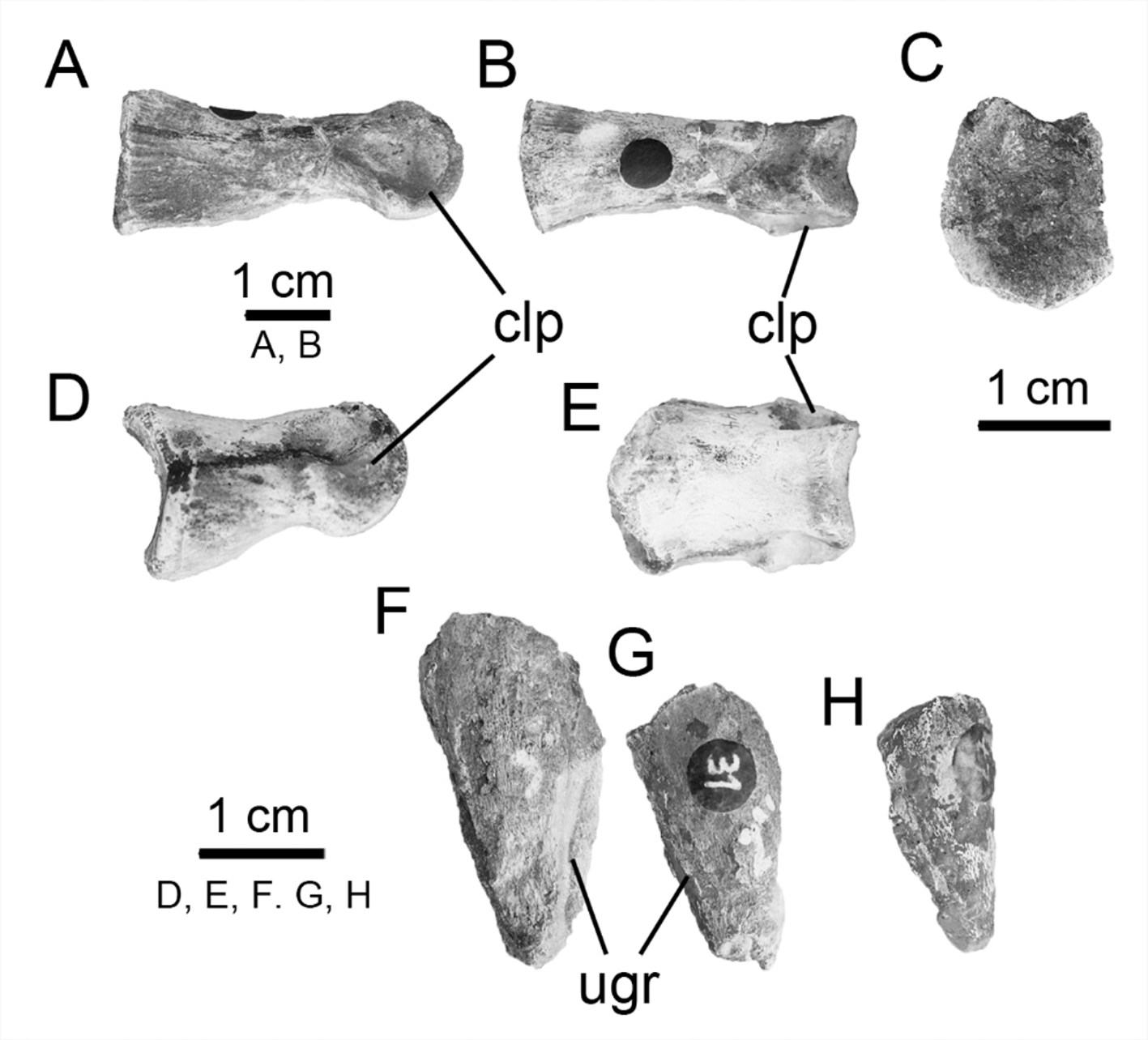
Non-ungual phalanx. One of the best-preserved proximal non-ungual phalanges ( Fig. 10 View FIGURE 10 A–B–C), probably the first phalanx, exhibits a concave surface for articulation with the metatarsal. The shaft is slightly constricted as in Gasparinisaura . The distal articulation of the phalanx possesses two almost symmetrical condyles. These are separated by a marked intercondylar groove. The phalanx has ovoid collateral ligament pits on the external surfaces of the lateral and medial condyles (although the latter is not as strongly developed as the former). Another well preserved non-ungual phalanx ( Fig. 10 View FIGURE 10 D–E; a second phalanx) is shorter than the previously described phalanx. The proximal articular surface is divided by a marked sagittal ridge (= intercondylar ridge, according to Zheng et al. 2012). This strong dorsal process delimits two concave surfaces. This suggests that it is not a proximal phalanx [(based on anatomical comparisons with well represented pedes; e.g., Changchunsaurus and Yueosaurus), probably phalanx?-2]. The well-developed collateral ligament pits and the presence of dorsal and sagittal ridges on the proximal end of the non-proximal phalanx are features that are common to all basal neornithischians and basal ornithopods ( Zheng et al. 2012).
Ungual phalanges. The ungual phalanges are subtriangular in shape ( Fig. 10 View FIGURE 10 F–G–H). They bear welldeveloped grooves on both the lateral and medial sides and the ventral surface is planar. The elongate grooves extend for almost the full length of the unguals. The morphology of these elements resembles to those described in MOZ-PV 0 20 ( Canudo et al. 2013). In contrast, those seen in styracosternans are dorsoventrally flattened and spade-shaped.

| UNPSJB-Pv |
Universidad Nacional de la Patagonia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.