
Lathomicrus, Jałoszyński, Paweł, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.196159 |
|
DOI |
https://doi.org/10.5281/zenodo.6206102 |
|
persistent identifier |
https://treatment.plazi.org/id/6D5D87B9-FFFF-FFEB-A4F1-0A72FB5FCE55 |
|
treatment provided by |
Plazi |
|
scientific name |
Lathomicrus |
| status |
gen. nov. |
Genus Lathomicrus View in CoL gen. n.
Type species. Neseuthia sumatrana Franz, 1984 .
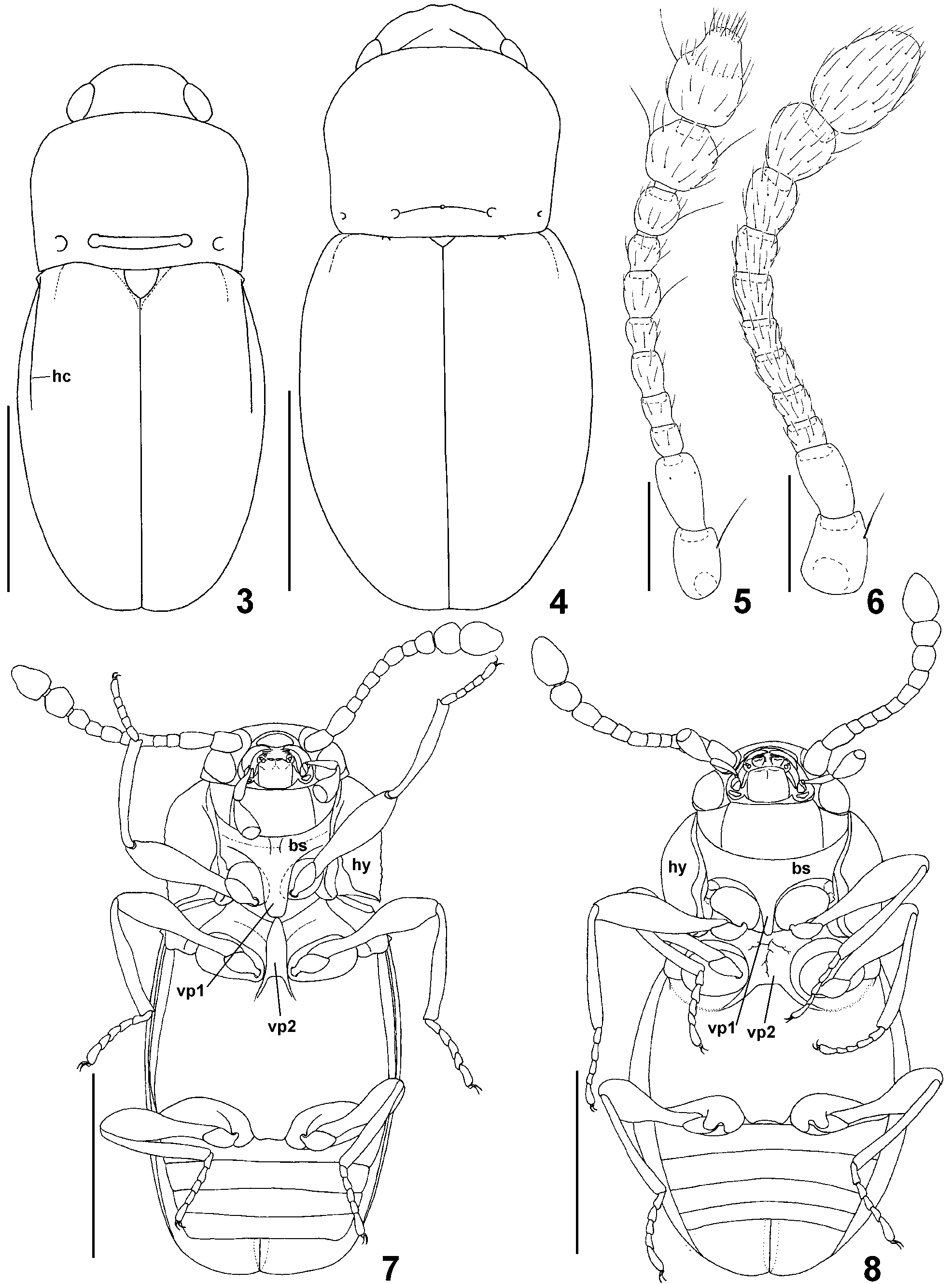
Diagnosis. Tempora distinct, short; maxillary palpomere III inversely conical, maxillary palpomere IV as broad as apex of palpomere III, button-shaped; pronotum with four ante-basal foveae, internal foveae connected by transverse groove; each elytron with humeral denticle connected with humeral carina, without subhumeral carina; prosternal intercoxal process with broadened ventral surface so that internal margins of procoxae in ventral view concealed by lateral margins of process ( Fig. 7 View FIGURES 3 – 8 ); mesoventral intercoxal process nearly equal in width to prosternal process, with subtriangular anterior part and divergent posterior arms ( Fig. 7 View FIGURES 3 – 8 ).
Description. General morphology ( Figs. 1 View FIGURES 1 – 2 , 3 View FIGURES 3 – 8 ) resembling Cephennomicrus ; body very small and slender, covered with distinct vestiture.
Head broadest at large eyes, with very short but distinct tempora; maxillary palps with very short palpomere I, strongly elongate and slightly clavate palpomere II, large and very broad palpomere IV strongly and gradually broadened from base to apex, and broad but very short palpomere IV; antennae with large scape and pedicell, slender and moderately compactly assembled antennomeres III–VIII and indistinctly delimited, three-segmented club, with terminal antennomere divided into cylindrical basal part and subconical apical part delimited by ring of setae. Antennae ( Fig. 5 View FIGURES 3 – 8 ) covered with sparse short and suberect setae and additionally with several long strongly erect setae located on antennomeres I, VII and IX–XI.
Pronotum ( Fig. 3 View FIGURES 3 – 8 ) subrectangular in shape, with rounded anterior margin, microserrate lateral margins forming entire lateral carinae, and bisinuate posterior margin; base with two lateral pairs of small foveae, internal foveae connected by transverse groove; margins of pronotum without long setae. Prosternum ( Fig. 7 View FIGURES 3 – 8 ) with short basisternal part; prosternal intercoxal process broad and with laterally expanded ventral surface (Tshaped in cross-section), so that internal margins of procoxae in ventral view are concealed under lateral margins of prosternal process; hypomera broad, not divided.
Mesoventrite ( Fig. 7 View FIGURES 3 – 8 ) with mesoventral intercoxal process comparable in width to procoxal process, lateral margins of process are nearly parallel in middle, convergent anteriorly to form subtriangular shape, and divergent posteriorly to form distinct posterior arms.
Elytra ( Figs. 1 View FIGURES 1 – 2 , 3 View FIGURES 3 – 8 ) entire, oval, each with small humeral denticle connected posteriorly to long longitudinal carina; basal fovea indistinct. Mesoscutellum subtriangular, well visible in intact specimen.
Metaventrite ( Fig. 7 View FIGURES 3 – 8 ) long, with metaventral intercoxal process broader than mesoventral process; mesepisterna visible in ventral view.
Legs ( Fig. 7 View FIGURES 3 – 8 ). Procoxae short, nearly spherical; mesocoxae oval; metacoxae strongly transverse; all femora clavate; tibiae and tarsi slender.
Abdomen with six visible, non-modified sternites.
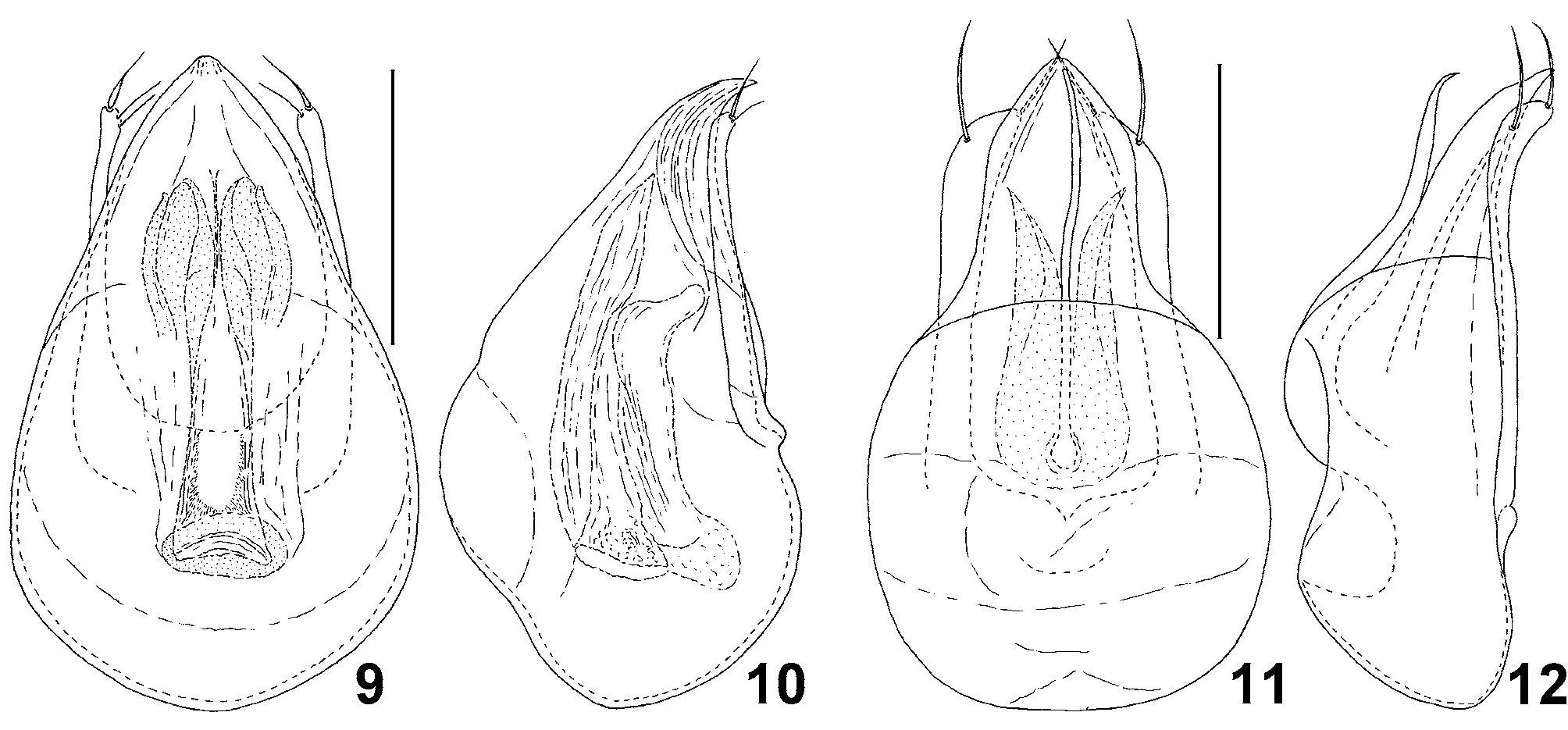
Aedeagus ( Figs. 9, 10 View FIGURES 9 – 12 ) symmetrical, with tubular internal armature; parameres free, with apical setae.
Distribution. Sumatra.
Etymology. The name is derived from a Greek noun "λάθOς" (lathos), meaning "escape from detection" (contemporarily also "an error, fault, mistake, oversight"), combined with the Greek adjective "μɭκρός" (mikros) by analogy to Cephennomicrus and to emphasize the extremely small body size of the type species. Gender masculine.
Remarks. The new genus shares a number of characters with other Cephenniini , especially with Cephennomicrus, Cephennula Jałoszyński, 2008 b and Trurlia Jałoszyński, 2009a, all of them possibly forming a distinct lineage within the tribe. Members of this complex differ from all other genera of Cephenniini in having more than two ante-basal foveae on the pronotum (a single pair or lack of pronotal foveae in all other cephenniines). Lathomicrus shows several characters that may be interpreted as transitions between structures found in Cephennomicrus and Cephennula. In Cephennomicrus the elytra lack longitudinal humeral carinae, which are present in Lathomicrus and Cephennula, but the latter has additionally subhumeral carinae. The internal pair of ante-basal pronotal foveae in some species of Cephennomicrus are connected by a transverse groove, in some others the groove is missing, while the foveae in Cephennula and Lathomicrus are connected. The prosternal intercoxal process in Cephennomicrus and Lathomicrus lacks the anterior margin (i.e., its anterior part is not separated from the basisternum), while in Cephennula and Trurlia it is separated from the basisternum and has a clearly delimited anterior part. However, in Cephennomicrus the prosternal process is more or less rectangular in cross-section, so that the internal margins of procoxae are visible in ventral view ( Fig. 8 View FIGURES 3 – 8 ), whereas in Lathomicrus , Cephennula and Trurlia the prosternal process has laterally expanded ventral surface, so that it is T-shaped in cross-section and the internal margins of procoxae are not visible in ventral view ( Fig. 7 View FIGURES 3 – 8 ). Therefore, the prosternal intercoxal process of Lathomicrus represents a transition between the simple process in Cephennomicrus and the strongly expanded process in Cephennula and Trurlia. The mesoventral intercoxal process of Lathomicrus may also be regarded as a transitional form between Cephennomicrus and Cephennula, with an additional step in the morphocline represented by Trurlia. In Cephennomicrus the mesoventral process is not delimited from the ventrite anteriorly and is typically very broad ( Fig. 8 View FIGURES 3 – 8 ) and relatively low (i.e., only slightly protruding ventrally), but shows a tendency to reducing in width in some species, e.g. in C. minimus ( Franz, 1983) (illustrated in Jałoszyński 2009b). In Trurlia, the process is narrow and higher (i.e., more protruding ventrally), but still not delimited anteriorly. In Lathomicrus the mesoventral process is narrow, higher (more expanded ventrally) than in Cephennomicrus and Trurlia, clearly delimited anteriorly, and in cross-section subrectangular making the internal margins of mesocoxae well visible in ventral view ( Fig. 7 View FIGURES 3 – 8 ). In Cephennula the mesoventral process is the most derived; it is not only high, narrow and delimited anteriorly, but also its ventral surface is expanded in a way similar to the prosternal process, so that the internal margins of mesocoxae are also hidden and not visible in ventral view.
Genera of the Old World Cephenniini can be identified by means of the key presented below. The remaining, exclusively Neotropical genera require revision and will be treated separately.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.