
Phrynobatrachus horsti, Rödel, Mark-Oliver, Burger, Marius, Zassi-Boulou, Ange Ghislain, Emmrich, Mike, Penner, Johannes & Barej, Michael F., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4032.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:5368E4BD-2AA0-46C6-B532-A60DE6CFDA57 |
|
DOI |
https://doi.org/10.5281/zenodo.6096751 |
|
persistent identifier |
https://treatment.plazi.org/id/6F7D87C9-FFD9-FFBB-E7CF-DCB835CEF82E |
|
treatment provided by |
Plazi |
|
scientific name |
Phrynobatrachus horsti |
| status |
sp. nov. |
Phrynobatrachus horsti View in CoL sp. nov.
Figs. 1–2 View FIGURE 1 View FIGURE 2
Holotype. ZMB 81698 (adult male; field # MBUR 3237), Republic of the Congo, Niari Department, 02°12’47.8’’ S, 12°50’06.0’’ E, 28 September 2012, leg. M. Burger & A.G. Zassi-Boulou.
Paratypes (all from Republic of the Congo, all collected by M. Burger & A.G. Zassi-Boulou). ZMB 81699 (adult male; field # MBUR 3098), Kouilou Department, Yombo River, 04°23’26.1’’ S, 11°40’26.0’’ E, 27 July 2012; ZMB 81700 (adult male; field # MBUR 3122), ZMB 81701 (adult female; field # MBUR 3123), Kouilou Department, Yombo River, 04°23’26.1’’ S, 11°40’26.0’’ E, 29 July 2012; ZMB 81702 (adult male; field # MBUR 3148), ZMB 81703 (adult female; field # MBUR 3149), Kouilou Department, Yombo River, 04°23’26.1’’ S, 11°40’26.0’’ E, 31 July 2012; ZMB 81704 (adult female; field # MBUR 3236), Niari Department, 02°12’47.8’’ S, 12°50’06.0’’ E, 28 September 2012; ZMB 81705 (adult male; field # MBUR 3286), Niari Department, 02°12’47.8’’ S, 12°50’06.0’’ E, 30 September 2012; ZMB 81706 (adult male; field # MBUR 3357), Niari Department, 02°12’47.8’’ S, 12°50’06.0’’ E, 4 October 2012; ZMB 81707 (adult female; field # MBUR 3541), Niari Department, 02°19’36.2’’ S, 12°47’32.7’’ E, 26 October 2013; ZMB 81708 (adult male; field # MBUR 3591), Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 19 October 2013; ZMB 81709 (adult female; field # MBUR 3610), ZMB 81710 (adult male; field # MBUR 03611), Niari Department, 02°19’36.2’’ S, 12°47’32.7’’ E, 21 October 2013; ZMB 81711 (adult female; field # MBUR 3612), ZMB 81712 (adult male; field # MBUR 3613), Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 20 October 2013; ZMB 81713 (adult male; field # MBUR 3641), Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 21 October 2013; ZMB 81714 (adult female; field # MBUR 3634), ZMB 81715 (adult male; field # MBUR 3652), ZMB 81716 (adult female; field # MBUR 3653), ZMB 81717 (adult female; field # MBUR 3677), ZMB 81718 (adult male; field # MBUR 3678), Niari Department, 02°19’36.2’’ S, 12°47’32.7’’ E, 26 October 2013; ZMB 81719 (adult male; field # MBUR 3681), ZMB 81720 (adult male; field # MBUR 3682), ZMB 81721 (adult male; field # MBUR 3683), Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 23 October 2013; ZMB 81722 (adult female; field # MBUR 3696), ZMB 81723 (adult female; field # MBUR 3697), ZMB 81724 (adult male; field # MBUR 3698), ZMB 81725 (adult male; field # MBUR 3699), ZMB 81726 (adult female; field # MBUR 3732), ZMB 81727 (adult male; field # MBUR 3742), Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 23 October 2013; ZMB 81728 (adult male; field # MBUR 3770), ZMB 81729 (adult male; field # MBUR 3771), Niari Department, 02°24’08.8’’ S, 12°59’08.0’’ E, 14 November 2013; ZMB 81730 (adult male; field # MBUR 4005), ZMB 81731 (adult male; field # MBUR 4006), Niari Department, 02°24’08.8’’ S, 12°59’08.0’’ E, 26 November 2013.
Additional material deposited in IRSEN (all from the Republic of the Congo, all collected by M. Burger & A.G. Zassi-Boulou). MBUR 3028–3031, Kouilou Department, Yombo River, 04°23’26.1’’ S, 11°40’26.0’’ E, 22 July 1012; MBUR 3038–3039, Kouilou Department, Yombo River, 04°23’33.1’’ S, 11°40’16.0’’ E, 21 July 2012; MBUR 3063, 3099, 3103–3106, Kouilou Department, Yombo River, 04°23’26.1’’ S, 11°40’26.0’’ E, 23, 26 and 27 July 2012; MBUR 3641, Niari Department, 02°12’38.3’’ S, 12°49’56.6’’ E, 21 October 2013; MBUR 4005, Niari Department, 02°24’08.8’’ S, 12°59’08.0’’ E, 26 November 2013.
Diagnosis. The genetic data (16S rRNA) identifies the new species as a member of the genus Phrynobatrachus . Morphologically this is confirmed by: the general body shape; the presence of a tarsal tubercle; and the presence of pedal webbing. From the morphologically similar Arthroleptis they differ by lacking a median dorsal skin raphe and a black tympanic spot. From juvenile Ptychadena they can be distinguished by the lack of parallel dorsal ridges.
Compared to other Phrynobatrachus the new species can be identified by the combination of the following characters: having snout-vent lengths smaller than 20 mm; males lacking tooth like projections in the lower jaw; males having black throats with spines; lacking a spiny eyelid tubercle; possessing a distinct lateral black facial mask; lacking X-shaped dorsal ridges; possessing comma-shaped dorsal ridges; lacking distinct markings on the whitish coloured belly; having yellowish ventral colouration on hind limbs; showing reduced pedal webbing and small but distinct discs on fingers and toes; and by a unique 16S rRNA sequence.
Differential diagnosis. The new species differs from other western African Phrynobatrachus species by: a small compact body (<20 mm; adults larger than 20 mm: P. acridoides , P. africanus , P. auritus , P. batesii , P. brongersmai , P. cornutus , P. cricogaster , P. danko , P. francisci , P. giorgii , P. intermedius , P. jimzimkusi , P. latifrons , P. liberiensis , P. plicatus , P. natalensis , P. njiomock , P. sandersoni , P. schioetzi , P. steindachneri ); lacking tooth like projections in lower jaw of males (present in P. africanus and P. sandersoni ); a black lateral face mask originating from at least the level of eyes and extending to below the supratympanic ridge (black face mask only shared with: P. auritus , P. batesii , P. chukuchuku , P. intermedius , P. plicatus , P. ruthbeateae ; less distinct brown face masks: P. werneri ; some P. rainerguentheri have entirely black heads); small, but distinct tympanum (tympanum absent according to original description: P. ogoensis ; indistinct but visible in BMNH 1947.2.6.83); black axillary spots of varying size, but generally being small and irregular shaped (larger distinct axillary and/or lateral spots and bands: P. dispar , P. fraterculus , P. gutturosus , P. maculiventris , P. pintoi , P. ruthbeateae ); males with a black throat (yellow in P. acridoides , P. alleni , P. batesii , P. fraterculus , P. latifrons ; throat with large black or brown spots: P. brongersmai , P. cornutus , P. giorgii ; white or whitish greyish: P. annulatus , P. dispar , P. hieroglyphicus ), the entire males’ throat—but in particular the centre—beset with black spines (black spines on throat shared with: P. chukuchuku , P. danko , P. jimzimkusi , P. ruthbeateae , P. steindachneri ; spines hyaline or white: P. hylaios , P. schioetzi ; P. werneri ; spines mostly along throat margins: P. leveleve , P. danko ); belly of males dirty white, dark grey towards flanks (belly at least partly yellow in P. alleni , P. batesii ; P. manengoubensis , P. werneri ; orange to yellow: P. danko ; black: P. chukuchuku ; white or bluish with black pattern: P. villiersi ; white with blackish spots or dots: P. annulatus , P. ghanensis , P. hylaios , P. maculiventris , P. pintoi ; a black circle and median spot on white or yellow background—bull’s eye pattern: P. cricogaster ); females with yellowish grey throat, belly dirty greyish (throat and belly with brown blotches: P. intermedius , P. jimzimkusi , P. njiomock , P. steindachneri ); lower parts of hind limbs in both sexes uniform yellow (yellow colouration shared with: P. alleni , P. batesii , P. danko , P. manengoubensis , possibly also with P. sternfeldi ; greyish/brownish to yellowish in P. ruthbeateae , P. schioetzi ; vaguely dark blotched in P. hylaios ); absence of spiny eyelid tubercles (spiny eyelid tubercle present in P. annulatus , P. calcaratus , P. cornutus , P. pintoi , P. taiensis , P. villiersi ); comma-shaped scapular ridges converging towards vent (scapular ridges long and converging towards mid-body, almost X-shaped: P. alleni , P. auritus , P. brongersmai , P. jimzimkusi , P. plicatus , P. steindachneri ; scapular ridges indistinct or absent: P. hieroglyphicus , P. intermedius , P. liberiensis , P. maculiventris , P. manengoubensis , P. tokba ); short dorsolateral ridges often distinct (short dorsolateral ridges shared with: P. r u t h be a t e a e); dorsal skin granular (smooth: P. fraterculus , P. hieroglyphicus , P. maculiventris , most P. rainerguentheri , P. tokba ; tuberculate: P. annulatus , P. calcaratus , P. cornutus , P. cricogaster , P. guineensis , P. sternfeldi , most P. guttursous , P. jimzimkusi , P. steindachneri , P. sternfeldi , P. taiensis , P. villiersi ; the new species described below); ventral skin smooth in both sexes (ventral skin with asperities in both sexes: P. chukuchuku ); absence of a blue band along the interior half of ventral shanks (present in P. auritus ); absence of nuptial pads in males (nuptial pads or swollen thumb in males present: P. b at e s i i, P. e l b e r t i, P. guineensis , P. hieroglyphicus , P. maculiventris , P. ogoensis , P. werneri ); distinct roundish discs on finger and toe tips (discs absent: most other species e.g. P. hieroglyphicus , P. intermedius , P. liberiensis , P. maculiventris , P. ogoensis , P. plicatus , P. rainerguentheri , P. tokba , P. werneri ; discs heart-shaped: P. sandersoni ); carpal gland in males lacking (carpal gland present: P. africanus ); femoral glands absent (present in males: P. calcaratus , P. cornutus , P. fraterculus , P. leveleve , P. maculiventris , P. ogoensis , P. phyllophilus ; present in both sexes: P. sandersoni ); rudimentary but distinct webbing (webbing indistinct or absent: P. annulatus , P. calcaratus , P. chukuchuku , P. cornutus , P. dispar , P. fraterculus , P. gutturosus , P. hieroglyphicus , P. maculiventris , P. manengoubensis , P. tokba , P. villiersi ; webbing more developed: P. acridoides , P. africanus , P. alleni , P. auritus , P. brongersmai , P. cricogaster , P. danko , P. elberti , P. francisci , P. giorgii , P. intermedius , P. jimzimkusi , P. latifrons , P. liberiensis , P. natalensis , P. njiomock , P. ogoensis , P. plicatus , P. steindachneri , P. sternfeldi ); small round tarsal tubercle (numerous tarsal tubercles fused to tarsal ridge: P. cricogaster ).
Morphologically the new species can be only mistaken for P. ruthbeateae and P. ba t es i i. A detailed comparison with these two taxa is presented in the discussion. A comparison with the second new species described herein, is included in the diagnosis of the latter species. For further character states of western African Phrynobatrachus species see: Peters (1870); Boulenger (1906a, b); Nieden (1910); Sternfeld (1917; Arthroleptis taeniatus ; replacement name by Ahl 1924: A. sternfeldi ; a Phrynobatrachus species according to Frétey 2008); de Witte (1921); Parker (1936); Angel (1940); Perret (1957, 1959, 1966); Guibé (1959); Guibé & Lamotte (1963); Lamotte & Xavier (1966a, b); Rödel (2000); Rödel & Ernst (2002b); Rödel et al. (2005, 2009a, b, 2010, 2011, 2012a, b); Uyeda et al. (2007); Zimkus (2009); Blackburn (2010); Blackburn & Rödel (2011); and Zimkus & Gvoždík (2012).
Description of holotype [measures in mm]. Typical, small adult male Phrynobatrachus with short oval, compact body shape; snout-vent length: 14; short snout, rounded to slightly truncate in dorsal and rounded to slightly protruding in lateral view; canthus rostralis rounded; loreal region straight to very slightly convex; headwidth directly behind the eyes: 5; medium sized eyes positioned laterally; eye-diameter: 2.1; distance anterior corner of eye to nostril: 1.5; nares small and roundish to slightly vertically elliptical, positioned laterally; straight distance nostril-snout tip: 1.1; nostril closer to snout than to eye; inter-narial distance: 2.4, larger than inter-orbital distance: 1.8; pupil round; supratympanic ridge curving down from posterior corner of eye to forearm base; tympanum diameter: 1.1, with distinct tympanic annulus, annulus round with central round papilla; tympanum much smaller than diameter of eye; upper and lower maxillae and praemaxillae with minute teeth, hidden by lips; vomerine teeth absent; elongated, almost rectangular tongue, tip with distinct v-shaped notch, median conical papilla near anterior attachment of tongue; tongue densely beset with small round papillae; small round choanae positioned at lateral edges of mouth roof.
Forelimbs slender; upper arm: 3.4; lower arm: 3.6; hand to tip of finger III: 4.6; hand with two large tubercles, small and oval thenar tubercle, large and round palmar tubercle; carpal gland absent; fingers with round, nondivided, protruding subarticular tubercles, no further tubercles on hand and fingers; relative finger length: IV<I=II<III; manual webbing absent; finger tips broadened, forming small roundish to slightly pointed discs, being almost double as wide as subarticular tubercles; no distinct nuptial pads; hind limbs short and slender; femur: 7.1, slightly shorter than tibia: 8.5; tibia length 60.7% of SVL; foot including longest toe: 10.3; femoral glands absent; tarsal tubercle present, small and round; large inner metatarsal tubercle: 0.8, long and narrow, approximately 1/3 of length of toe I: 2.8; small round outer metatarsal tubercle; long and slender toes; relative toe length: I<II<V<III<IV; digits with basal webbing; webbing formula I(1), II(1), III(2), IV(3), V(2); lateral skin fringes extending further towards toe tips; toe tips enlarged to roundish discs, being approximately double as wide as subarticular tubercles.
Dorsal skin fine granular (in life) to smooth (in preservation); a pair of long comma-shaped scapular ridges, ranging from about posterior corner of upper eyelid to slightly beyond level of forearm bases, converging towards vent; dorsolateral ridges faintly indicated; posterior edge of eyelid with few flat round warts, spiny eyelid tubercle absent; no white spines or warts on flanks; skin on outer parts of thighs and dorsal part of lower legs granular; skin on lower parts of limbs, belly and pectoral region smooth; throat without skin folds running parallel to mandibles; anterior third of lower mandible and anterior third of throat densely beset with rounded spines.
Colouration in ethanol after 1.5 years. Back and dorsal part of flanks more or less uniform brown; snout slightly clearer, dorsal with dark median spot, snout tip including anterior part of upper lip and loreal region light brown; clear spot between anterior part of eyelids; posterior part of inter-orbital space with dark bar; eyelids dark grey; lateral part of upper lip, sub-orbital region, supratympanic ridge and temporal area including tympanum dark brown to black, extending into a similarly coloured blotch on the anterior part of the upper arm; flanks posterior to forearm bases with large dark axillary blotch; brown flanks fainting ventrally into whitish belly; arms coloured as back, lower arm towards hand with two dark transversal bars; hands dorsally lighter coloured with dark crossbands; lower legs brown as back; thighs with 3-4 faintly indicated dark cross-bars; anterior part of thigh with dark longitudinal stripe, being the border to whitish ventral parts; lower leg dorsally with three (right) and four (left) dark cross-bars; feet and toes with dark cross-bands; dark blotch around vent, dorsally bordered by narrow yellowish line; lower parts of feet and hands greyish brown; remaining ventral parts of limbs and posterior part of belly uniform whitish; lateroventral parts of venter, anterior part of belly and pectoral region whitish but densely beset with minute black spots; throat and lower mandibles dark grey to black; lower mandibles with few small clear spots.
Colouration in life ( Fig. 1 View FIGURE 1 ). Basic colour of dorsal surface of head, upper part of flank, back and dorsal surfaces of extremities light brown; snout, loreal region and anterior part of eyelids orange brown; snout with median longitudinal black spot; black inter-orbital bar; back with irregular darker patterning; lower arms, thighs, lower legs and feet with dark crossbars; anterior part of upper mandible brown with a dark bar; a V-shaped darker figure in scapular region, pointing anteriorly; black lateral face mask extending from below the eye and supratympanic ridge to forearm bases, covering tympanum; continuing below arms as a lateroventral black patch to axillar region; eye posterodorsally bordered by fine blue line; black pupil; iris copper red; central belly dirty white; belly laterally and pectoral area dark grey, no distinct black ventrolateral line or band; ventral part of forelimbs and hands dark grey; groin area and lower surfaces of thighs, lower leg and tarsus yellowish to orange; lower part of lower legs bordered by blackish spots; ventral surfaces of toes blackish.
Variation ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ). Snout-vent length in males with spiny black throats (presumably indicating maturity) 13.5–15.8 mm and 13.1–19.7 mm in females (smallest one presumably not adult); body dimensions generally as in holotype, for small deviations see Table 1 View TABLE 1 . The scapular ridges may differ in lengths and distinctness, but are always discernible and comma-shaped. Short dorsolateral ridges, composed of a very dense row of small warts, are visible in most specimens. Scapular and dorsolateral ridges are usually coloured dark. The webbing formulae showed some slight variation ( Table 1 View TABLE 1 ). The black face mask may cover either the entire lateral parts of the head (sometimes with minute white spots; Figs. 2 View FIGURE 2 c, d), extending from snout-tip to dorsal of the forearm insertion, or the parts anterior to the eye are coloured in clearer brown ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), sometimes with darker spots. Individuals with clearer coloured loreal regions also have dark brown upper mandible with black bars instead of entirely black mandibles. The back might be uniform dark or reddish brown, show some darker patterning, or carry a broad central dark band ( Fig. 2 View FIGURE 2 a). A light inter-orbital stripe is usually present. The iris colour varies from golden to copper red. The flank might be either uniform brown, grey or mottled with darker dots and lines ( Fig. 2 View FIGURE 2 c). The black axillary spot may be distinct and extending as far caudally as to about mid-body. Usually, the edges of this spot are irregularly shaped. In other individuals it is nothing but a rather indistinct mottling of blackish spots with clearer dots. We neither observed in males, nor in females distinct black bordering of belly and lower shanks as in P. ruthbeateae . The inguinal region is yellowish. The belly seems to be dirty white in males and females (in preserved specimens it appears pure white); the border of the belly being dark grey in males. In some males, all with distinct gular spines, only the anterior half of the throat is coloured black. Females with dirty white (life) or white
(in preservation) throats. Their lower mandibles largely coloured black. This black colour often interrupted along mandibles and always interrupted at tip of mouth; the margins of black colour to throat skin are irregular, almost wavelike. Only indistinctly indicated lateral skin folds running parallel to mandible in males. Female throat skin smooth.
Genetics. The mean genetic difference in the sampled region of the 16S rRNA gene between Phrynobatrachus horsti sp. nov. and 40 other West African and Central African congeners ranged from 3.66–18.10% ( Table 2 View TABLE 2 ). The species with highest genetic similarity to P. horsti sp. nov. all occur in Central Africa. With the exception of P. ruthbeateae and P. b at e s i i, all belong to a clade of species being endemic to restricted mountainous regions along the Cameroon Volcanic Line (Zimkus et al. 2010; Zimkus & Gvoždík 2013). The uncorrected pairwise sequence divergence to the most similar species were P. ruthbeateae (mean: 3.66%), P. chukuchuku (3.78%), P. schioetzi (3.86%), P. steindachneri (3.89%), P. w er n e r i (4.09%), P. n j i om oc k (4.22%), P. jimzimkusi (4.28%), P. danko (4.43%), P. manengoubensis (4.48%), and P. b at e s i i (4.70%), respectively. A comparison of our sequences with the published sequences for Gabonese “ P. ruthbeateae ” records ( Zimkus & Larson 2013a, b; see Table 2 View TABLE 2 ), revealed that these frogs are conspecific with P. horsti sp. nov. The intraspecific variation of all available P. horsti sp. nov. sequences ranged from 0–1.56% (mean ± sd: 0.51 ± 0.43%; N= 105). Mean intraspecific variation in the morphologically similar P. ruthbeateae was 0.66% (0–1.20%, sd: 0.47, N= 6).
Acoustics ( Fig. 4 View FIGURE 4 ). The advertisement call of Phrynobatrachus horsti sp. nov. consists of numerous short single pulses, ranging from 7–10 ms (mean ± sd, 8.7 ± 1.1 ms, N= 337). The entire call has a duration of 2.1–4.5 sec (3.0 ± 0.9 sec, N= 12), with a pulse rate of 7.6–12.8 pulses/sec (9.4 ± 1.9 pulses/sec, N= 12). The calls start with a longer interval between the first and second pulse (15.1–529.9 ms; mean ± sd: 271.8 ± 181.3 ms, N= 12). These intervals then become shorter (16.6–188.7 ms; 67.1 ± 46.5 ms, N= 12). The dominant frequency of the calls range between 3161–3960 Hz, with the maximum call energy at 3656 Hz (N=12). The frequency range covers 2640 Hz (N=12), with a minimum frequency at 2177 Hz and a maxium frequeny at 4817 Hz. We observed neither harmonics nor any frequency modulation. Call parameters are summarized in Table 3 View TABLE 3 .
Distribution. Phrynobatrachus horsti sp. nov. is known from several localities across south-western Republic of the Congo (Kouilou and Niari Departments) and adjacent Gabon ( Fig. 4 View FIGURE 4 ). The latter locality has been published as a record of the morphologically similar P. ruthbeateae . However, the photos and genetics of the Gabon individuals ( Zimkus & Larson 2013a, b), confirm that these frogs are P. horsti sp. nov. (Appendix 1, Table 2 View TABLE 2 ). In contrast, it is not clear if the small puddle frogs termed Phrynobatrachus sp. 2 by Burger et al. (2004), originating from mid-altitude forests on the Monts Doudou, Gabon, are P. ruthbeateae , P. horsti sp. nov. or a third species. So far it seems that the new species may have a distribution in Central African lowland to mid-altitude rainforests inbetween the Ogooué and the Congo Rivers.
Habitat and natural history. Our P. horsti sp. nov. records stem from leaf litter in disturbed and undisturbed lowland to mid-altitude rainforests, including swamp forests ( Fig. 5 View FIGURE 5 ). The altitudinal range of P. horsti covers altitudes from 554–728 m a.s.l. in the Mayoko area, but drops below 100 m in the Kouilou Department. The species seems to breed in small and shallow stagnant pools, including holes excavated by artisanal gold miners. We heard many calling males at puddles in a logged forest patch on 20 and 21 October 2012. Three males and a female were found active on the forest floor near the edge of a stream on 26 July 2012. This is comparable to the habitat choice and calling sites of the closely related P. r u t h be a t e a e ( Rödel et al. 2012a). It is not entirely clear in which type of habitat P. horsti was recorded on the near-by Batéké Plateau in Gabon (400–840 m a.s.l.) by Zimkus & Larson (2013a, b). From the site descriptions it seemed to be a mix of savannah and gallery forest. In their table these authors mark the species for savannah, farmbush, gallery forest and forest ( Zimkus & Larson 2013b). A range of habitats from savannah to forest is unusual for the genus. Most other congeners show clear species specific habitat preferences (e.g. Rödel & Ernst 2002b; Rödel et al. 2004; Channing & Howell 2006; Kouamé et al. 2008). Comparable to us, Zimkus & Larson (2013b) state that they collected the frogs from leaf litter.
Etymology. The specific epithet ‘horsti’ is a patronym. The first author dedicates this species to his father, Horst Rödel. From earliest childhood he supported MOR’s fascination for nature in general and for “lower” vertebrates in particular. The suggested English name for this new species is Horst’s Puddle Frog.


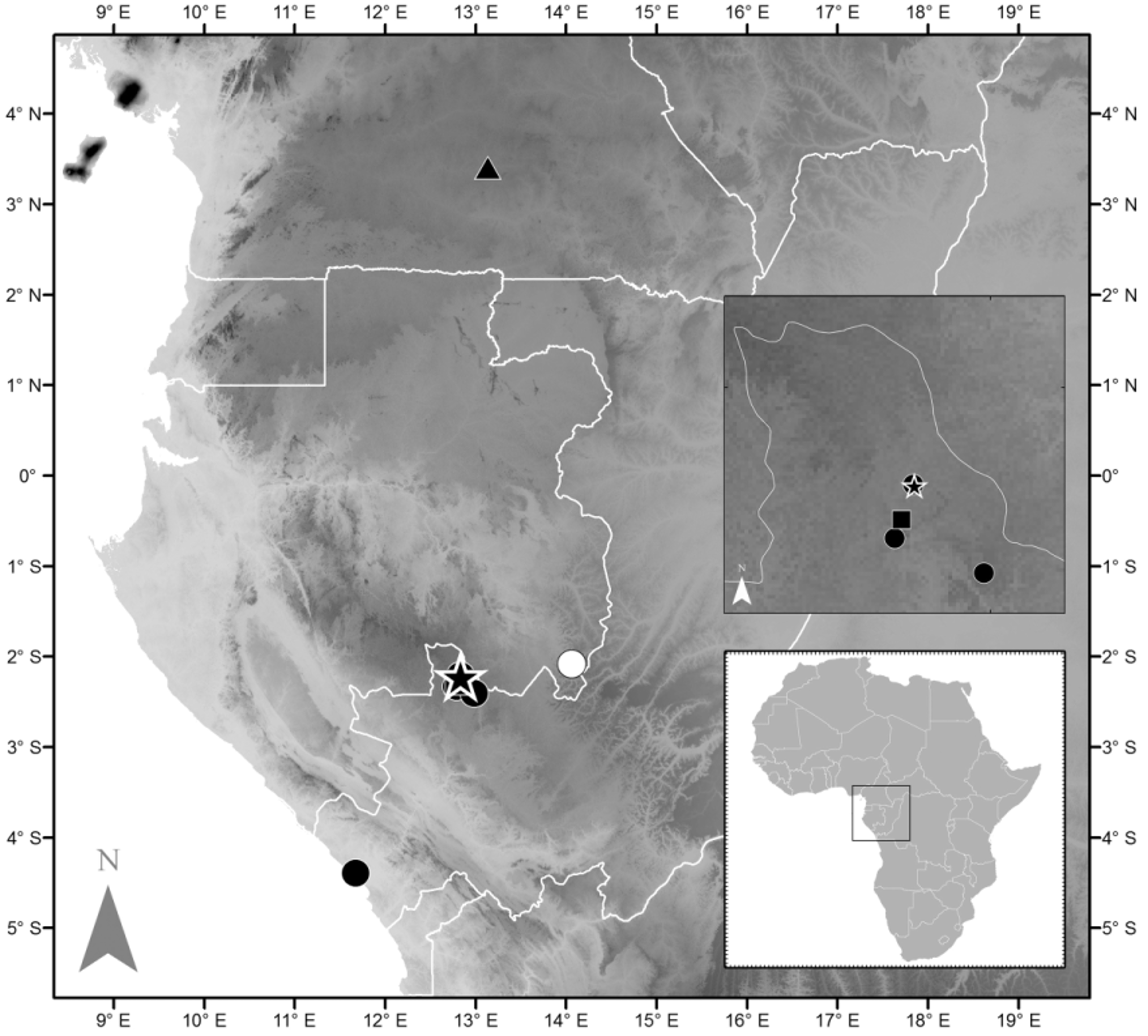

TABLE 1. Measurements [mm] of the type series of Phrynobatrachus horsti sp. nov. (holotype in bold); m = male, f = female, SVL = snout-vent length, HW = head width, ED = horizontal eye diameter; IO = inter-orbital distance (minimum distance between eyelids); IN = inter-narial distance; DEN = distance from anterior corner of eye to nose, DNS = straight distance from nose to snout-tip; TD = horizontal tympanum diameter; FL = femur length, TL = tibia length; FLT = foot length including toe IV; IMT = length of inner metatarsal tubercle, webbing formulae according to Rödel (2000); gaps = respective measure could not be assessed.
| ZMB# sex | SVL | HW | ED | IO | IN | DEN | DNS | TD | FL | TL | FLT | IMT | Webbing formulae |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 81698 m | 14.0 | 5.0 | 2.1 | 1.8 | 2.4 | 1.5 | 1.1 | 1.1 | 7.1 | 8.5 | 10.3 | 0.8 | I(1), II(1), III(2), IV(3), V(2) |
| 81699 m | 14.2 | 5.7 | 2.4 | 1.9 | 1.9 | 1.6 | 1.1 | 0.8 | 8.0 | 8.7 | 11.5 | 0.7 | I(1), II(1), III(2), IV(3), V(2) |
| 81700 m | 14.7 | 5.5 | 2.4 | 2.2 | 2.6 | 1.8 | 1.1 | 1.0 | 8.3 | 8.7 | 13.0 | 1.2 | I(1), II(1), III(2), IV(3), V(2) |
| 81701 f | 18.4 | 6.8 | 2.3 | 2.2 | 2.3 | 1.8 | 1.4 | 1.1 | 10 | 10.1 | 14.0 | 1.0 | I(1), II(1), III(2-1.75), IV(3), V(1.5) |
| 81702 m | 14.2 | 5.5 | 2.0 | 2.4 | 2.3 | 1.4 | 1 | 0.9 | 8.5 | 9.5 | 12.0 | 1.0 | I(1), II(1), III(2), IV(3), V(2) |
| 81703 f | 19.1 | 5.5 | 2.5 | 2.2 | 2.2 | 1.9 | 1.1 | 1.4 | 9.5 | 10.3 | 13.4 | 1.0 | I(1), II(1), III(2), IV(3), V(2) |
| 81704 f | 18.1 | 5.7 | 2.3 | 2.0 | 2.2 | 1.7 | 1.2 | 1.0 | 8.8 | 8.4 | 12.1 | 1.1 | I(1), II(1), III(2), IV(3), V(2) |
| 81705 m | 13.9 | 4.1 | 1.8 | 1.5 | 1.9 | 1.5 | 1.0 | 1.3 | 7.5 | 8.0 | 10.4 | 0.8 | I(1), II(1), III(2), IV(3), V(2) |
| 81706 m | 14.9 | 5.3 | 2.2 | 2.0 | 2.4 | 1.4 | 1.0 | 1.0 | 7.0 | 7.1 | 10.1 | 0.8 | I(1), II(1), III(2), IV(3), V(2) |
| 81707 f | 18.3 | 6.0 | 2.2 | 2.5 | 2.2 | 2.0 | 1.0 | 1.0 | 8.4 | 8.0 | 12.1 | 1.0 | I(1), II(1), III(2), IV(3), V(2) |
| 81708 m | 15.2 | 5.4 | 2.0 | 1.9 | 2.2 | 1.5 | 1.4 | 1.1 | 7.8 | 7.5 | 1.0 | I(1), II(1), III(2), IV(3), V(2) | |
| 81709 f | 16.0 | 5.5 | 2.3 | 1.9 | 2.0 | 1.5 | 1.1 | 1.2 | 8.1 | 8.4 | 10.1 | 0.8 | I(1), II(1), III(2), IV(2.75), V(1) |
| 81710 m | 15.2 | 4.1 | 1.9 | 2.1 | 2.1 | 1.5 | 1.4 | 1.1 | 7.1 | 7.4 | 10.8 | 0.8 | I(1), II(1), III(2.5-2), IV(3), V(2) |
| 81711 f | 17.4 | 6.2 | 2.4 | 1.5 | 2.0 | 1.4 | 1.2 | 1.4 | 8.1 | 9.1 | 13.4 | 0.9 | I(1), II(1), III(2), IV(3), V(2) |
| 81712 m | 13.7 | 4.0 | 1.9 | 2.1 | 2.0 | 1.4 | 1.1 | 1.1 | 7.0 | 7.0 | 10.9 | 0.7 | I(1), II(1), III(2), IV(3), V(2) |
| 81713 m | 15.2 | 4.3 | 1.8 | 1.8 | 1.8 | 1.6 | 1.2 | 1.0 | 8.1 | 7.6 | 12.0 | 1.0 | |
| 81714 f | 18.1 | 6.1 | 2.5 | 2.3 | 2.3 | 2.0 | 1.4 | 1.0 | 8.6 | 9.4 | 13.4 | 1.1 | I(1), II(1), III(2), IV(3), V(2) |
| 81715 m | 14.1 | 4.0 | 1.9 | 2.0 | 1.9 | 1.5 | 1.1 | 1.0 | 7.4 | 7.9 | 11.4 | 0.8 | |
| 81716 f | 17.5 | 5.7 | 2.2 | 1.9 | 1.8 | 1.7 | 1.2 | 1.3 | 9.9 | 9.5 | 14.0 | 0.8 | I(1), II(1.5-1), III(2), IV(3), V(2) |
| 81717 f | 13.9 | 5.1 | 2.2 | 2.1 | 2.2 | 1.4 | 1.1 | 1.2 | 7.3 | 8.4 | 11.4 | 0.7 | I(1), II(1), III(2), IV(3), V(2) |
| 81718 m | 15.5 | 4.1 | 1.9 | 2.1 | 1.8 | 1.1 | 1.0 | 1.1 | 7.2 | 8.2 | 11.8 | 0.8 | I(1), II(1), III(2), IV(3), V(2) |
| 81719 m | 15.8 | 5.5 | 2.5 | 2.2 | 2.2 | 1.4 | 1.2 | 1.0 | 7.4 | 7.9 | 10.2 | 0.9 | I(1), II(1), III(2), IV(3), V(2) |
| 81720 m | 13.9 | 4.2 | 2.1 | 2.2 | 1.9 | 1.4 | 1.0 | 1.2 | 7.8 | 0.8 | I(1), II(1), III(2), IV(3), V(2) | ||
| 81721 m | 13.9 | 4.3 | 1.9 | 2.1 | 2.1 | 1.5 | 1.0 | 1.0 | 8.2 | 8.3 | 0.9 | ||
| 81722 f | 19.1 | 5.3 | 2.2 | 2.6 | 1.8 | 1.9 | 1.5 | 1.2 | 9.0 | 9.8 | 9.4 | 0.9 | I(1), II(1), III(2), IV(3), V(2) |
| 81723 f | 17.5 | 6.3 | 2.3 | 2.1 | 2.3 | 1.8 | 1.0 | 1.5 | 8.9 | 9.1 | 12.5 | 0.9 | I(1), II(1), III(2), IV(3), V(2) |
| 81724 m | 15.1 | 5.4 | 2.1 | 2.5 | 2.1 | 1.4 | 1.1 | 1.3 | 8.1 | 8.1 | 10.2 | 1.0 | I(1), II(1), III(2), IV(3), V(2) |
| 81725 m | 14.4 | 5.1 | 2.1 | 2.1 | 1.8 | 1.5 | 1.1 | 1.0 | 7 | 7.4 | 7.6 | 0.7 | I(1), II(1), III(2), IV(3), V(2) |
| 81726 f | 16.3 | 6.0 | 2.2 | 1.8 | 2.0 | 1.6 | 1.0 | 1.0 | 8.1 | 8.8 | 12.2 | 0.7 | I(1), II(1), III(2), IV(3), V(2) |
| 81727 m | 15.5 | 5.4 | 2.0 | 1.9 | 2.0 | 1.4 | 1.1 | 1.0 | 8.1 | 8.1 | 11.5 | 0.8 | I(1), II(1), III(2), IV(3), V(2) |
| 81728 m | 13.5 | 5.5 | 1.9 | 2.3 | 2.3 | 1.4 | 1.3 | 1.2 | 7.4 | 8.1 | 10.1 | ||
| 81729 m | 14.8 | 4.1 | 2.1 | 2.2 | 2.0 | 1.8 | 1.0 | 1.1 | 7.5 | 8.5 | 10.1 | ||
| 81730 m | 15.5 | 4.7 | 2.1 | 1.8 | 2.1 | 1.4 | 1.2 | 1.0 | 8.2 | 7.8 | |||
| 81731 m | 14.3 | 5.1 | 2.0 | 2.1 | 2.1 | 1.5 | 1.1 | 1.2 | 7.3 | 8.8 | 10.9 | 1.0 | I(1), II(1), III(2), IV(2.75), V(1) |
TABLE 2. Genetic similarity of 15 Phrynobatrachus horsti sp. nov. with 40 other West and western Central African Phrynobatrachus species. Given are minimum (min), maximum (max) and mean values of uncorrected pairwise sequence divergence, presented in % of 348 – 515 bp of the 16 S rRNA gene; sd = standard deviation; N = number of individual inter-specific comparisons; for museum accession and corresponding GenBank accession numbers see Appendix 1; * = compare text.
| min | max | mean | sd | N | |
|---|---|---|---|---|---|
| acridoides | 10.27 | 11.78 | 10.70 | 0.33 | 15 |
| africanus | 8.66 | 9.85 | 9.03 | 0.28 | 15 |
| alleni | 13.05 | 14.08 | 13.23 | 0.27 | 15 |
| annulatus | 11.93 | 12.53 | 12.33 | 0.15 | 15 |
| auritus | 11.07 | 12.46 | 11.34 | 0.33 | 15 |
| batesii | 4.31 | 4.97 | 4.70 | 0.18 | 15 |
| calcaratus | 11.93 | 13.16 | 12.37 | 0.26 | 15 |
| chukuchuku | 3.22 | 4.23 | 3.78 | 0.23 | 15 |
| cornutus | 11.73 | 12.35 | 12.00 | 0.16 | 15 |
| cricogaster | 4.84 | 5.45 | 5.09 | 0.16 | 15 |
| danko | 4.04 | 4.85 | 4.43 | 0.20 | 15 |
| dispar | 11.67 | 12.34 | 12.08 | 0.19 | 15 |
| francisci | 10.06 | 11.40 | 10.36 | 0.32 | 15 |
| fraterculus | 11.55 | 11.96 | 11.75 | 0.12 | 15 |
| ghanensis | 12.80 | 14.41 | 13.08 | 0.40 | 15 |
| guineensis | 12.01 | 13.36 | 12.29 | 0.33 | 15 |
| gutturosus | 12.32 | 13.86 | 12.92 | 0.32 | 15 |
| hylaios | 15.30 | 17.83 | 15.75 | 0.59 | 15 |
| intermedius | 8.27 | 9.17 | 8.70 | 0.22 | 15 |
| jimzimkusi | 3.82 | 4.92 | 4.28 | 0.25 | 15 |
| latifrons | 13.76 | 15.56 | 14.03 | 0.46 | 15 |
| leveleve | 10.75 | 11.55 | 11.16 | 0.19 | 15 |
| liberiensis | 6.93 | 8.40 | 7.43 | 0.32 | 15 |
| maculiventris | 13.30 | 14.26 | 13.84 | 0.23 | 15 |
| manengoubensis | 4.22 | 4.88 | 4.48 | 0.17 | 15 |
| mayokoensis sp. nov. | 11.70 | 12.72 | 12.11 | 0.21 | 30 |
| natalensis | 10.89 | 12.11 | 11.40 | 0.26 | 15 |
| njiomock | 3.62 | 4.63 | 4.22 | 0.23 | 15 |
| cf. ogoensis * | 12.22 | 13.31 | 12.65 | 0.23 | 15 |
| phyllophilus | 11.43 | 12.04 | 11.74 | 0.16 | 15 |
| pintoi | 13.72 | 15.01 | 14.19 | 0.27 | 15 |
| plicatus | 10.70 | 12.52 | 11.10 | 0.42 | 15 |
| rainerguentheri | 11.97 | 12.99 | 12.12 | 0.27 | 15 |
| ruthbeateae | 2.62 | 4.74 | 3.66 | 0.44 | 60 |
| sandersoni | 17.57 | 18.50 | 18.10 | 0.23 | 15 |
| schioetzi | 3.42 | 4.67 | 3.86 | 0.30 | 15 |
| steindachneri | 3.43 | 4.44 | 3.89 | 0.22 | 15 |
| tokba | 10.12 | 11.62 | 10.75 | 0.30 | 15 |
| villiersi | 12.33 | 13.60 | 12.95 | 0.26 | 15 |
| werneri | 3.83 | 4.44 | 4.09 | 0.16 | 15 |
TABLE 3. Advertisement call parameters of Phrynobatrachus horsti sp. nov. Given are: call number (call #); dominate frequency (DoFr) in Hz; minimum frequency (minFr) in Hz; maximum frequency (maxFr) in Hz; frequency bandwidth (FrBW) in Hz; pulse number (Pu #); pulse rate (PuRa) per second; pulse duration (PuDu) in milliseconds; pulse interval at the beginning (PuInS) and at the end of a call (PuInE) both in milliseconds; and call duration (CD) in seconds. For all call characteristics, minimum, maximum, mean and standard deviation values are provided.
| Call# | DoFr | minFr | maxFr | FrBW | Pu# | PuRa | PuDu | PuInS | PuInE | CD |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3852.7 | 2282.0 | 4510.0 | 2227.0 | 37 | 11.6 | 9.2 | 235.9 | 82.5 | 3.2 |
| 2 | 3161.1 | 2093.0 | 4699.0 | 2606.0 | 20 | 7.9 | 10.3 | 240.6 | 103.9 | 2.5 |
| 3 | 3823.2 | 1912.0 | 5435.0 | 3523.0 | 35 | 9.2 | 7.1 | 415.1 | 51.4 | 3.8 |
| 4 | 3486.6 | 2098.0 | 5398.0 | 3300.0 | 28 | 9.6 | 8.2 | 237.5 | 52.8 | 2.9 |
| 5 | 3725.7 | 1986.0 | 5583.0 | 3597.0 | 36 | 7.9 | 8.0 | 465.5 | 30.4 | 4.5 |
| 6 | 3644.5 | 1875.0 | 5212.0 | 3337.0 | 32 | 7.6 | 8.2 | 529.9 | 69.1 | 4.2 |
| 7 | 3513.3 | 1986.0 | 5324.0 | 3337.0 | 33 | 8.4 | 7.9 | 363.5 | 16.6 | 3.9 |
| 8 | 3658.7 | 2172.0 | 4397.0 | 2225.0 | 26 | 12.6 | 10.0 | 15.1 | 68.3 | 2.1 |
| 9 | 3692.7 | 2283.0 | 4359.0 | 2077.0 | 24 | 8.8 | 8.1 | 266.5 | 25.0 | 2.7 |
| 10 | 3959.6 | 2839.0 | 4359.0 | 1520.0 | 17 | 8.0 | 8.6 | 22.3 | 188.7 | 2.1 |
| 11 | 3707.2 | 2320.0 | 4285.0 | 1965.0 | 19 | 8.7 | 8.4 | 451.7 | 82.1 | 2.2 |
| 12 | 3641.7 | 2283.0 | 4248.0 | 1965.0 | 30 | 12.8 | 10.8 | 17.8 | 34.8 | 2.3 |
| Min | 3161.1 | 1875.0 | 4248.0 | 1520.0 | 17.0 | 7.6 | 7.1 | 15.1 | 16.6 | 2.1 |
| Max | 3959.6 | 2839.0 | 5583.0 | 3597.0 | 37.0 | 12.8 | 10.8 | 529.9 | 188.7 | 4.5 |
| Mean | 3655.6 | 2177.4 | 4817.4 | 2639.9 | 28.1 | 9.4 | 8.7 | 271.8 | 67.1 | 3.0 |
| SD | 204.9 | 259.4 | 524.9 | 734.2 | 6.9 | 1.9 | 1.1 | 181.3 | 46.5 | 0.9 |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.