
Bokermannohyla pseudopseudis (Miranda-Ribeiro, 1937)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4527.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:F8149959-4B98-433A-9982-629AAA01F0BB |
|
DOI |
https://doi.org/10.5281/zenodo.5967663 |
|
persistent identifier |
https://treatment.plazi.org/id/705E4058-FFD5-FF8F-57F5-4A6E8062FEEA |
|
treatment provided by |
Plazi |
|
scientific name |
Bokermannohyla pseudopseudis |
| status |
|
Bokermannohyla pseudopseudis View in CoL
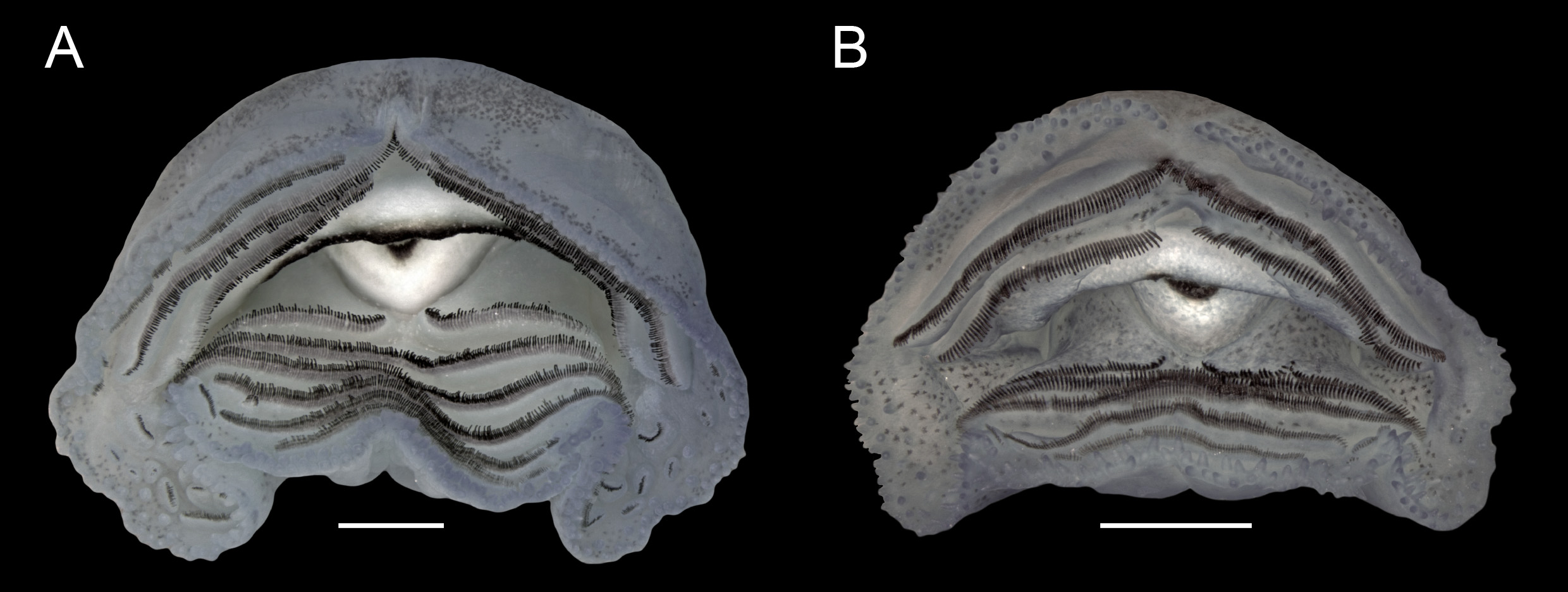
External morphology (individuals from Alto Paraíso de Goiás; Table 1). Body depressed (BH/BW = 0.74–0.80;
Fig. 1A, B View FIGURE 1 ), BL 0.33–0.36 times TL; elongated-elliptical in dorsal view; in lateral view, ventral contour flat in peribranchial region, slightly convex in abdominal region. Snout rounded in dorsal (BWN/BWE = 0.71–0.79) and lateral views. Nostrils small (ND/BL = 0.02–0.03), elliptical, dorsally positioned (IND/BWN = 0.47–0.53), dorsolaterally directed, located halfway between eyes and tip of snout (NSD/ESD = 0.45–0.50); poorly developed fleshy projection present on medial margin. Eyes medium-sized (ED/BWE = 0.18–0.21), dorsally located (IOD/ BWE = 0.66–0.71), dorsolaterally directed. Spiracle sinistral, lateral, visible in dorsal and ventral views (SVD/BH = 0.40–0.60), short (SL/BL = 0.08–0.14), posterodorsally projected; inner wall with distal portion free from body and longer than external wall; opening elliptical, slightly narrower than anterior portion of spiracular tube, located at medial third of the body (SSD/BL = 0.59–0.65). Vent tube short (VTL/BL = 0.11–0.16), dextral ( Fig. 1C View FIGURE 1 ); ventral wall fused to ventral fin and slightly longer than dorsal wall. Tail moderately high (MTH/TAL = 0.33– 0.36), higher than body (MTH/BH = 1.22–1.52); tail musculature robust (TMH/BH = 0.70–0.83) reaching tip of pointed tail. Dorsal and ventral fins moderately high (DFH/TAL = 0.10–0.13; VFH/TAL = 0.07–0.09), with convex external margins; dorsal fin originating on posterior third of body at a low slope (DFiA = 5°–20°); maximum height at middle third of tail; ventral fin origin concealed by vent tube. Oral disc ( Fig. 2A View FIGURE 2 ) ventrally positioned (ODP = 11°–18°), large (ODW/BW = 0.41–0.54, measured with oral disc folded); posterior margin with three emarginations (one medial and two lateral); marginal papillae alternate, rounded and conical, arranged mostly in a single row, or small biserial regions, interrupted anteriorly by a narrow gap (AGL/ODW = 0.06–0.07; Fig. 2A View FIGURE 2 ). Many submarginal papillae (18–22) scattered laterally in angular region. Labial tooth row formula 3(Ae1, 2)/6(1); gaps in Ae1, A2 and P1 corresponding to 25, 20 and 7% of oral disc width, respectively; anteriormost and posteriormost rows with smaller teeth than those of other rows. Several flaps with labial teeth (about 20 in each angular region) located laterally in oral disc. Jaw sheaths highly dekeratinized, with only distal margins darkcolored; lateral processes of upper jaw sheath laterally directed. Lateral line system evident in life and in preserved specimens. In dorsal view, supraorbital line with 16–22 stitches running from posterior region of eyes and converging anteriorly towards nares and diverging towards tip of snout. Posterior infraorbital line with 4–7 stitches. Posterior supraorbital line with 4–6 stitches located near posterior region of supraorbital line. Two lines extending from mid-body posteriorly throughout tail length: dorsal line with 8–10 sparse stitches located medially, converging before body-tail junction and continuing posteriorly with 7–15 stitches along dorsal fin base until middle third of tail; middle-body line with 19–22 stitches located laterally, joining middle caudal series with 37–43 stitches. Middle caudal series curving upward dorsally along approximate middle third of tail, reaching dorsal fin base. In lateral view, infraorbital line with 25–28 stitches beginning in posterodorsal region of eyes, circling eyes ventrally, and extending slightly curved towards the snout until anterior region of oral disc. Anterior oral line with about 20 stitches extending from lateral region of oral disc to venter, approaching angular line. Angular line with 19–21 stitches descending vertically from eyes until longitudinal oral line, where it curves abruptly and posteriorly towards the venter. Longitudinal oral line with 6–8 stitches, extending towards angular line, parallel to infraorbital line. Ventral body-line with 28–32 stitches, extending from near vent tube to above spiracle; after two gaps without neuromasts at spiracle level, ventral body-line continuing posteroventrally until mid-abdominal region.
Coloration. In life, body dark brown to grayish, marbled, commonly with large rounded black blotches scattered mainly dorsally, which reach the tail muscle in some specimens. Spiracle edge lightly pigmented ( Fig. 3A, B View FIGURE 3 ). Ventral body varies in color from silver to lead. Ventral translucent skin highlights the well-developed m. rectus abdominis. Tail musculature light brown, gradually darkening along its length. Fins cream-colored with reddish flecks, marbled by rounded dark brown and golden blotches especially distributed posteriorly. Well-defined whitish spots (e.g., Kolenc et al. 2008; Pezzuti et al. 2015) anterolateral to the base of the vent tube, commonly above stitches of the ventral line; seven specimens also with 1–16 similar whitish spots scattered on the venter. In preservative, body coloration like that in life, but paler ( Fig. 1 View FIGURE 1 ). Old preserved individuals lose pigmentation with the color varying from light brown or gray to cream. Tail fins become transparent.
Variation. Individuals at Stage 25 exhibited considerable variation in size, reaching a maximum of 80.1 mm ( Table 1). Three specimens at Stage 25 (UFMG 1972) had fewer (4–8) submarginal papillae. Two specimens at Stage 25 had only three flaps with labial teeth laterally in the oral disc. Three specimens at Stage 25 (UFMG 1972) had seven posterior rows of labial teeth. Some individuals did not possess rounded black blotches on the dorsal region of the body. Tadpoles from Cavalcante (UFMG 1973-75) were very similar to those from the type locality, only differing by having slightly larger eyes (ED/BWE = 0.21–0.23). All specimens from Cavalcante had LTRF 3(Ae1, 2)/6(1), and dekeratinized jaw sheaths.
Natural history notes. Bokermannohyla pseudopseudis tadpoles are benthic. They are found throughout the year in well-preserved permanent rocky streams and rivulets associated with natural grasslands or narrow riparian forests with open canopies. Individuals can be found in large backwater pools with slow-flowing water, located immediately below waterfalls ( Fig. 3E View FIGURE 3 ). The pools have quartzitic beds, and some are more than three meters deep; the larvae are found from the shore to the deepest parts of the pools. The tadpoles are crepuscular/nocturnal and are frequently found scratching biofilm and algae from rock surfaces. Although some individuals are observed by day, most take shelter between rocks, dead leaves or macrophytes. Syntopic tadpoles included Ameerega flavopicta (Lutz) ; Boana ericae (Caramaschi & Cruz) ; B. lundii (Burmeister) ; Odontophrynus salvatori Caramaschi ; Proceratophrys goiana (Miranda-Ribeiro) ; Scinax rupestris Araujo-Vieira, Brandão , & Faria; and Ololygon skaios (Pombal, Carvalho, Canelas, & Bastos).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.