
Plecotus macrobullaris ( Kuzjakin, 1965 )
|
publication ID |
https://doi.org/ 10.1093/mspecies/sey003 |
|
publication LSID |
lsid:zoobank.org:pub:CBE7455F-57F2-4CA7-A126-D4EF8F8C6D24 |
|
DOI |
https://doi.org/10.5281/zenodo.4592243 |
|
persistent identifier |
https://treatment.plazi.org/id/706E878D-FFFA-4A5A-57B0-FB9CFDD8FD0F |
|
treatment provided by |
Felipe |
|
scientific name |
Plecotus macrobullaris ( Kuzjakin, 1965 ) |
| status |
|
Plecotus macrobullaris ( Kuzjakin, 1965) View in CoL
Alpine Long-eared Bat
Plecotus auritus macrobullaris Kuzjakin, 1965:76 View in CoL . Type locality “near Vladikavikaz, North, Ossetia, Russ. Fed.”
Plecotus alpinus Kiefer and Veith, 2002:8 View in CoL . Type locality “Ristolas, Haute-Alpes, France.”
Plecotus microdontus Spitzenberger et al., 2002:5 . Type locality “Lienz, Eastern Tyrol, Austria.”
CONTEXT AND CONTENT. Order Chiroptera , family Vespertilionidae , subfamily Vespertilioninae , tribe Plecotini . Plecotus macrobullaris is 1 of the 11 Palearctic species recognized in the genus Plecotus ( Spitzenberger et al. 2006) . Several authors have suggested the presence of 2 geographically separated subspecies of P. macrobullaris ( Spitzenberger et al. 2003; Benda et al. 2004; Kiefer and von Helversen 2004): P. macrobullaris macrobullaris (Eastern group) and P. macrobullaris alpinus (Western group). This division is supported by the presence of 2 geographically separated mitochondrial lineages that probably diverged more than 1 million years ago ( Alberdi et al. 2015b). The average mitochondrial dissimilarity between specimens of the 2 lineages is 3.4%. However, there is no information about the genomic structure of populations based on nuclear DNA, thus subspecific differentiation cannot be entirely confirmed. Additionally, the craniometric differentiation does not agree with the phylogenetic dissimilarity, as individuals from the Caucasus (Western lineage) and Alps (Eastern lineage) highly resemble each other, whereas individuals from the Balkans, Turkey, and Syria (Eastern lineage) occupy morphospaces of their own ( Spitzenberger et al. 2006).
NOMENCLATURAL NOTES. Plecotus macrobullaris was first described in the northern slope of the Greater Caucasus (North Ossetia) by Kuzjakin in 1965, as a subspecies of Plecotus auritus , namely P. auritus macrobullaris ( Kuzjakin 1965) . In 2001– 2002, 2 research groups almost simultaneously reported the existence of a clade distinct from P. auritus and P. austriacus in the Austrian Alps ( Spitzenberger et al. 2001; Kiefer et al. 2002). Subsequently, each research group described it as a new species ( Kock 2002), assigning 2 different names: Plecotus alpinus (Kiefer and Veith 2002) and Plecotus microdontus ( Spitzenberger et al. 2002) . In 2003, molecular contrast between the newly described species and P. auritus macrobullaris described nearly 40 years earlier by Kuzjakin, indicated they belong to the same clade ( Spitzenberger et al. 2003). Therefore, the original nomenclature was recognized as the valid name for the species, but upgraded to the species level.
DIAGNOSIS
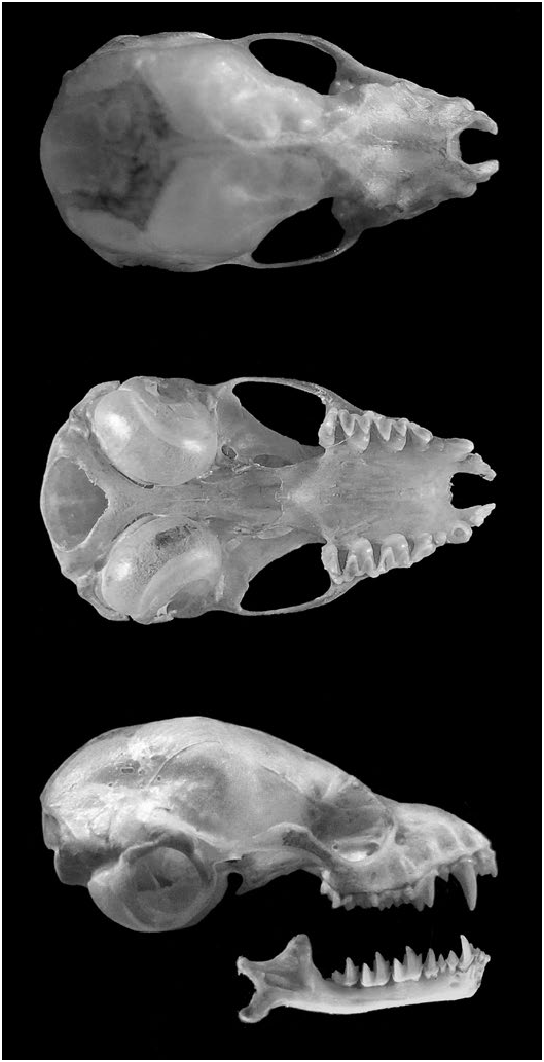
Plecotus macrobullaris ( Figs. 1 View Fig and 2 View Fig ) shares high morphological similarity with the other Plecotus species in areas where their distributions overlap, namely the brown longeared bat P. auritus , the gray long-eared bat P. austriacus , and Kolombatovic’s long-eared bat P. kolombatovici . The most conspicuous diagnostic characteristic of P. macrobullaris is the inverted triangular-shaped chin pad that most individuals (e.g., 95% in the Swiss Alps—Ashrafi et al. 2010) exhibit below the lower lip ( Spitzenberger et al. 2002). The chin pad of the other species tends to be rounded and is often darker in young individuals (Sachanowicz and Ciechanowski 2006). However, variation in color, size, and shape of the chin pad can lead to inconsistent identification. Male specimens can be identified with high reliability based on penis shape ( Ashrafi et al. 2010). P. macrobullaris specimens exhibit a cylindrical penis, whereas the penis of P. austriacus and P. kolombatovici males is club-shaped and the penis of P. auritus narrows toward the end (Dietz and von Helversen 2004; Tvrtkovic et al. 2005). Although several authors have proposed other means of identification based on external qualitative characters such as the color of the ventral fur or the presence of hair on the hind feet (Kiefer and Veith 2002; Dietz and von Helversen 2004), most of them have not been validated across the entire geographic range of the species. Despite the large morphometric overlap between the 4 western Palearctic long-eared bats ( Tvrtkovic et al. 2005; Ashrafi et al. 2010), a multilinear combination of 8 morphometric measurements yielded an identification success rate of 97.5% among Plecotus from Switzerland. In addition, several authors reported that the thumb length is a relatively good criterion to differentiate P. macrobullaris (> 6.2 mm) from P. austriacus and P. kolombatovici (<6.6 mm) where these 3 species cohabit ( Spitzenberger et al. 2002; Tvrtkovic et al. 2005). All the mentioned Plecotus species can be unambiguously discerned using short mitochondrial molecular markers, such as the cytochrome b or the control region ( Juste et al. 2004; Spitzenberger et al. 2006), as well as the standardized COI barcode region ( Agnarsson et al. 2011).
GENERAL CHARACTERS
Plecotus macrobullaris is a medium-sized (6–10 g, forearm length usually between 39 and 43 mm) Palearctic bat. It has the characteristic long ears of plecotine bats, which are fully erect and extended forward when echolocating and in flight, and folded and curled sideways when at rest. Ears and tragus are light brown, and eyes are fairly large and black. The face is dark brown, with a robust snout and a pinkish mandible where the inverted triangular-shaped pad usually stands out on the chin. The ventral fur is dense and usually looks whitish, often with pale yellowish highlights both sides of the neck and shoulders; the dorsal fur is typically pale gray. All hairs are bicolored in adult specimens, with a black base and lighter tips. There is no sexual dimorphism in fur coloration. The patagium is light brown and translucent, without fur. Coloration, morphometric measurements, and the length of the fur are subject to considerable geographical variation. Dental formula is i 2/3, c 1/1, p 2/3, m 3/3, total 36.
Mean external biometric measurements (mm; with range and n) were: forearm length 41.2 (35.5–44.8, 195); length of hind foot 7.6 (6.6–9.0, 177); tragus length 17.2 (14.2–20.0, 166); tragus width 5.8 (5.0–7.0, 157); thumb length 6.9 (5.5–7.9, 192); and claw length 2.6 (0.9–3.6, 83—Kiefer and Veith 2002; Garin et al. 2003; Trizio et al. 2005; Tvrtkovic et al. 2005; Ashrafi et al. 2010). Females are usually larger than males ( Tvrtkovic et al. 2005; Ashrafi et al. 2010).
Mean craniometric measurements (mm; with range and n) were: greatest length of skull 17.0 (15.9–18.3, 82); condylocanine length 15.1 (14.1–16.1, 70); condylobasal length 15.8 (14.7–16.9, 80); height of braincase 7.6 (7.1–8.1, 80); breadth of braincase 8.5 (7.7–9.3, 75); breadth of interorbital constriction 3.5 (3.1–3.9, 81); maximum bulla diameter 4.6 (4.2–4.9, 57); anterior palatal breadth 3.8 (3.4–4.2, 75); maxillary breadth 6.3 (5.9–6.7, 81); length of upper toothrow from 1st incisor to 3rd molar 6.7 (6.3–7.2, n = 51); length of upper toothrow from canine to 3rd molar 5.6 (5.3–6.3, 87—Kiefer and Veith 2002; Spitzenberger et al. 2003; Benda et al. 2004). The cranial and dental structures are similar across the entire geographic distribution of P. macrobullaris , but the skull size and the auditory bullae tend to be smaller in the Western clade than in the Eastern clade ( Spitzenberger et al. 2003).
DISTRIBUTION
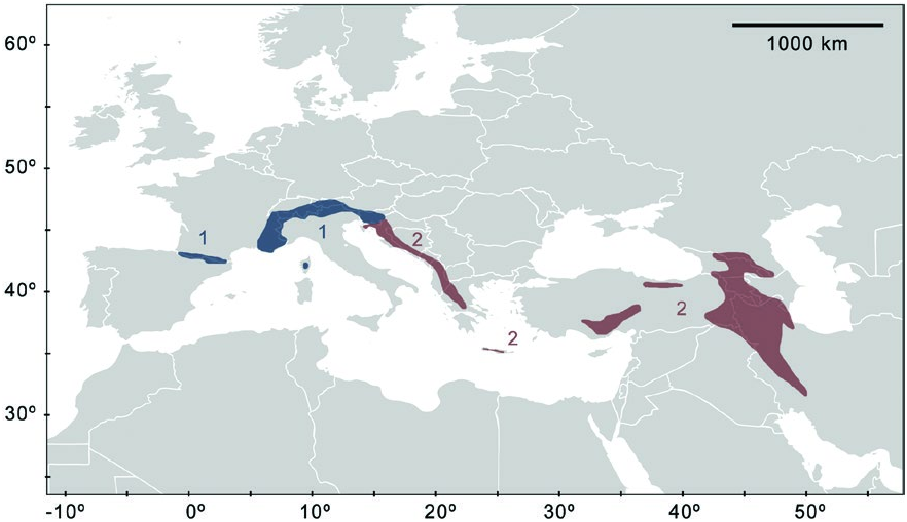
Plecotus macrobullaris is the only bat species with a palealpine distribution ( Fig. 3 View Fig ; i.e., widely distributed across the Palearctic, but limited to mountain environments—Alberdi et al. 2014). It is found in the main mountain ranges in the southwestern and central Palearctic, as well as the close surroundings ( Alberdi et al. 2013). To date, P. macrobullaris has been found in the Pyrenees ( France and Spain—Garin et al. 2003; Alberdi et al. 2013), Alps ( Austria, France, Italy, Liechtenstein, Slovenia, Switzerland—Chirichella et al. 2003; Kiefer and von Helversen 2004; Trizio et al. 2005; Spitzenberger et al. 2006; Presetnik et al. 2009; Mattei-Roesli 2010; Rutishauser et al. 2012), Dinaric Alps ( Albania, Bosnia-Herzegovina, Croatia, Macedonia, Montenegro, Serbia—Juste et al. 2004; Tvrtkovic et al. 2005; Sachanowicz and Ciechanowski 2006; Bekker and Boshamer 2007; Budinski et al. 2016), Pindos Mountains (Greece—Alberdi et al. 2013), several mountain ranges in Turkey (Kaçkar, Taurus, Guneydogu Toroslar—Karatas and Sozen 2006), Greater Caucasus ( Georgia, Russia—Spitzenberger et al. 2006; Kiefer 2008; Alberdi et al. 2013), Lower Caucasus ( Armenia, Azerbaijan—Kiefer 2008), Anti-Lebanon (Syria—Benda et al. 2006), and the Iranian Alborz and Zagros Mountains ( Benda et al. 2012). P. macrobullaris has been also found on 2 Mediterranean islands: Corsica and Crete (Kiefer and von Helversen 2004; Benda et al. 2008).
The elevational range of P. macrobullaris spans from sea level up to about 2,800 m, even though there are considerable differences between geographic areas. Almost all records in the Pyrenees and the Zagros mountains are above 1,500 m, but the species can be found at considerably lower elevations in very steep and mountainous regions. In such regions, P. macrobullaris shows an extremely wide elevational distribution (e.g., in the Alps it can be found between 200 and 2,200 m—Alberdi et al. 2013), and in Croatia from sea level up to mountain tops (Pavlinic and Tvrtkovic 2004). No fossils are known.
FORM AND FUNCTION
Echolocation calls are fairly similar to other Plecotus species. Plecotus macrobullaris emits multiharmonic, downward frequency-modulated signals consisting mostly of the 1st and the lower part of the 2nd harmonic ( Dietrich et al. 2006; Barataud 2014). The 1st harmonic starts at about 46 kHz and ends around 23 kHz, with a peak frequency around 35 kHz. The 2nd harmonic starts around 67 kHz and does not overlap with the 1st harmonic. The pulse length varies considerably depending on the surrounding environment, emitting short signals (up to 0.8 ms) when flying close to the clutter, and long signals (up to 7.3 ms) when flying far from the clutter. The longer the signal the lower the frequency band and the peak frequency. Although P. macrobullaris is genetically more closely related to P. auritus , its signal structure is closer to that of P. austriacus .
ECOLOGY
Plectous macrobullaris females aggregate in small groups (5–25) to form breeding colonies ( Ashrafi et al. 2010). These colonies may contain several nulliparous females, with an occasional presence of male bats ( Alberdi et al. 2015a).
Distribution records indicate that P. macrobullaris is ecologically linked to mountain environments (Kiefer and Veith 2002; Alberdi et al. 2013), although it is not restricted to high elevations (Pavlinic and Tvrtkovic 2004; Ashrafi et al. 2010). Using an ecological niche modeling approach, Alberdi et al. (2014) concluded that the broad-scale distribution of P. macrobullaris is mainly shaped by topographic factors, as it is found in areas with very steep topography but missing in flatlands. In contrast, the relative influence of climatic factors was much lower, as the distribution of P. macrobullaris covers a very broad climatic range, from the extremely cold and rainy Alps to the warm and dry mountains in the Middle East. Alberdi et al. (2014) suggested that its fondness for steep environments might be the result of increased availability of rocky and open-space areas that would provide appropriate foraging and roosting environments for P. macrobullaris .
However, the study of roosting and foraging preferences of P. macrobullaris has yielded contrasting results. Most of the known roosting sites of P. macrobullaris are man-made buildings ( Alberdi et al. 2013). In fact, it is one of the most common species found in church attics in the Alps ( Rutishauser et al. 2012). Nevertheless, results of a radiotracking study performed in the Pyrenees depicted a very different behavior. Alberdi et al. (2015a) captured 51 free-flying P. macrobullaris , and radiotracked them to their roosting sites. Most bats sheltered in rock crevices, and many males and nulliparous females made systematic use of scree deposits as roosts. Many of those roosting sites were found in alpine environments (above the upper forest treeline). Only 3 of the 51 tracked bats, all of them breeding females, roosted in buildings. The authors considered 2 possible explanations for the contrasting results. Bats (mainly breeding females) might make use of buildings more often in the Alps than in the Pyrenees because the climatic conditions are colder (about 2°C at the same elevation). Conversely, almost all the records in the Alps come from surveys of building that were considered suitable a priori, which entails a systemic bias toward man-made buildings. Similarly, the few roosts that were known in the Pyrenees before the radiotracking study were also buildings ( Alberdi et al. 2015a). Bats might also use natural roosting resources in the Alps, but the previously described approaches may not be suitable for revealing the entire roosting spectrum of P. macrobullaris .
Plecotus macrobullaris has been also found in caves, mostly in the southeastern area of its distribution ( Benda et al. 2008, 2012). Probably, it was originally linked to karstic areas with high availability of rock crevices, scree slopes, and caves. In fact, this preference to limestone environments can be still seen in the Pyrenees Mountains and Croatia ( Tvrtkovic et al. 2005; Alberdi et al. 2013). However, P. macrobullaris has been able to take advantage of relatively new roosting resources such as man-made buildings. Finally, a male P. macrobullaris was documented using tree hole indicating that they might occasionally use alternate types of roosts. These results depict a flexible roosting behavior, and suggest that depending on the environmental conditions, as well as sexual and reproductive status, P. macrobullaris bats may exhibit different roosting preferences ( Fig. 4 View Fig ).
Roosting-site habitat selection has also produced contrasting results. Rutishauser et al. (2012) reported that P. macrobullaris chooses buildings surrounded by forest environments, although Alberdi et al. (2015a) found that most roosts were located closer to meadows and open forests, and further away from deciduous forest and shrubbery. These conflicting results might reflect a geographical trend (i.e., bats roost closer to forest in the Alps than in the Pyrenees), or it might be due to the differences in the employed approaches and the roosting resources that were examined. Rutishauser et al. (2012) studied the location of manmade buildings where P. macrobullaris was found roosting, but Alberdi et al. (2015a) analyzed the roosts spotted after radiotracking free-ranging bats, mainly natural rock cracks and scree deposits. Interestingly, the building roosts identified in the latter study were located close to or surrounded by forest ( Alberdi et al. 2015a).
Regarding foraging preferences, the abundant captures performed hundreds of meters above the upper forest limit ( Alberdi et al. 2013), the roosts located far from forest environments ( Alberdi et al. 2015a), and the open-space moth species identified in its diet ( Alberdi et al. 2012) suggest that, at least in the Pyrenees, P. macrobullaris is an open-space forager. However, this pattern largely contrasts with the observations reported from the Swiss Alps, where Ashrafi et al. (2013) concluded that P. macrobullaris avoids grasslands and selects its foraging habitat according to the heterogeneity of vertical vegetation structures. Similarly, Pavlinic and Tvrtkovic (2004) reported that in Croatia the species appears linked to deciduous forests, but P. macrobullaris has never been observed in forests in the Pyrenees ( Alberdi et al. 2013). To further blur the picture of foraging preferences of P. macrobullaris , another radiotracking study carried out in the Italian Alps concluded that P. macrobullaris avoids woodlands ( Preatoni et al. 2011). Two radiotracking studies indicated that the foraging range of P. macrobullaris is usually below 4 km (Ashrafi 2010; Preatoni et al. 2011), although displacement of up to 7 km, and elevation differences of more than 1,000 m have been recorded in a single night ( Alberdi et al. 2015a).
The diet of P. macrobullaris is mainly composed of moths (Order Lepidoptera ), although it also consumes coleopterans and dipterans ( Ashrafi et al. 2011; Alberdi et al. 2012). Most moth species identified in its diet belong to the family Noctuidae , and 96% of prey species ( Alberdi et al. 2012) identified using molecular methods were tympanate moths (i.e., species that can detect bats’ echolocation calls—Goerlitz et al. 2010).
Multiple ecological and distribution data suggest the existence of competitive exclusion between P. macrobullaris and P. austriacus . Both species use similar open and semi-open areas for foraging, and their diets are basically composed of grassland moth species ( Razgour et al. 2011; Alberdi et al. 2012). Conversely, the trophic niche of P. auritus is different, as it also includes nonflying arthropods and is basically composed of woodland prey ( Ashrafi et al. 2011; Razgour et al. 2011). There are also large similarities in echolocation signals and flight behavior ( Dietrich et al. 2006). All these ecological resemblances likely yield a parapatric distribution of P. austriacus and P. macrobullaris in many geographic areas. In Switzerland P. austriacus occupies the Northern lowland areas, while P. macrobullaris is restricted to the mountainous regions ( Rutishauser et al. 2012). A similar geographical split is also observed in Croatia, where P. macrobullaris seems to be restricted to the karstic areas ( Tvrtkovic et al. 2005). In the southern slopes of the Pyrenees Mountains, the parapatric distribution responds to an altitudinal pattern rather than to a geographical one. In that mountain range, P. macrobullaris occupies the highest open areas and P. austriacus is found in lower zones ( Alberdi et al. 2013). There are also exceptions where both species can be found together, such as the Swiss city of Genève ( Rutishauser et al. 2012), or several subalpine areas in the Pyrenees ( Alberdi et al. 2013).
As occurs with the other Plecotus species, P. macrobullaris emits very faint echolocation pulses ( Dietrich et al. 2006). These pulses attenuate rapidly, making it almost impossible for most ultrasound detectors to record P. macrobullaris even from 5 m of distance. In addition, differentiation from other Plecotus species is also very challenging ( Barataud 2014). Consequently, its presence and abundance can be overlooked or underestimated in biodiversity surveys solely based on echolocation detection. Additionally, Alberdi et al. (2013) reported a surprisingly meager capture success of P. macrobullaris individuals in water points, suggesting that other capture techniques such as setting long netchains in open areas should be employed to increase capture efficiency and obtain a better picture of abundance of this species.
BEHAVIOR
When roosting in man-made buildings Plectous macrobullaris is usually found forming monospecific colonies, although it has been occasionally observed sharing attics with other Plecotus species; specifically P. kolombatovici and P. auritus ( Tvrtkovic et al. 2005; Ashrafi et al. 2010). In caves it has been found along with species of Rhinolophus and Myotis ( Benda et al. 2012) . Roost fidelity varies according to the type of roosting resource used, as well as the sex and reproductive condition of the individual. Individuals roosting in man-made buildings show very high roost fidelity and bats roosting in natural rock crevices exhibit slightly less fidelity. The lowest fidelity is found in the use of scree deposits, as these bats change their roosting location almost every day ( Alberdi et al. 2015a). Regarding sex and reproductive condition, breeding females barely change their roosting sites, nulliparous females show intermediate fidelity, although males exhibit a very labile behavior, switching their roosts almost every day ( Alberdi et al. 2015a).
There is no confirmed data about the hibernation behavior of P. macrobullaris . The only known information is based on 2 observations of Plecotus specimens hibernating in caves with ice formations above> 2,000 m in the Pyrenees Mountains and the Slovenian Alps ( Kryštufek 1987; Aihartza 1992). As both observations are prior to the description of P. macrobullaris , they were not identified as such. However, the knowledge on the ecology of Plecotus bats suggests that those observations most probably belong to P. macrobullaris .
GENETICS
Despite the morphological and ecological similarity to Plectous austriacus ( Ashrafi et al. 2011) , P. macrobullaris belongs to the so-called “auritus group” ( Juste et al. 2004; Spitzenberger et al. 2006). This group is divided into 2 main mitochondrial clades, between which 79.5% of the total genetic variation occurs ( Alberdi et al. 2015b). The presence of these 2 matrilineal lineages was reported by the 1st molecular analyses of P. macrobullaris ( Spitzenberger et al. 2003) , and was later confirmed using complete mitochondrial genomes ( Alberdi et al. 2015b). The 1st analyses based on short mitochondrial markers (e.g., control region) showed differences of about 2.5% between the 2 lineages ( Spitzenberger et al. 2003), although the complete mitochondrial sequences exhibited larger overall differentiation, namely 3.4% ( Alberdi et al. 2015b). All the individuals from the Pyrenees and the Alps belong to the Western clade ( Alberdi et al. 2015b), while the individuals from the other mountain ranges belong to the Eastern clade. Hence, the current contact zone involving the 2 lineages is located between the Eastern Alps and the Dinaric Alps (Eastern Italy, Slovenia, and Northern Croatia). All molecularly characterized individuals from Eastern Italy and Slovenia but 1 belongs to the Western clade, while all individuals from Croatia belong to the Eastern clade ( Spitzenberger et al. 2006; Alberdi et al. 2015b). Based on complete mitochondrial genomes, Alberdi et al. (2015b) reported that both lineages split more than 1 million year ago, before the beginning of the large Pleistocene climatic oscillations. Using paleoclimatic niche reconstructions the authors estimated that the Dinarides ( Croatia) remained ecologically unsuitable for most of the time during the Pleistocene, which could have acted as a natural ecological barrier that prevented the interbreeding between the lineages.
The genetic structure of both populations is rather different, reflecting different paleodemographic histories. The Eastern lineage conserves considerably larger genetic diversity than the Western lineage. The Eastern lineage began to diversify around 900,000 years ago and exhibits marked population structure. Conversely, the most recent common female ancestor of the Western lineage was estimated to live about 68,000 years ago, and diversification probably occurred after the last glaciation ( Alberdi et al. 2015b).
CONSERVATION
Plectous macrobullaris is listed as “Least Concern” by the International Union for Conservation of Nature and Natural Resources ( Hutson et al. 2016), but due to its recent discovery the actual conservation status of P. macrobullaris is still uncertain. However, the abundant captures performed in high-mountain environments ( Alberdi et al. 2013), as well as the large number of roosts found in buildings in the Alps ( Rutishauser et al. 2012) and the Dinarides (Pavlinic and Tvrtkovic 2004) suggest that it is currently a relatively abundant species in mountainous environments.
Although in the Pyrenees Mountains the species has been mainly found in natural roosts ( Alberdi et al. 2015a), most breeding colonies known in the Alps are in the attics of manmade structures, specifically churches ( Rutishauser et al. 2012). Hence, the conservation and management of such buildings are important for the conservation of the species. Because the Alps are one of the coldest regions in the geographic distribution of P. macrobullaris , the availability of warm shelters suitable for breeding, such as church attics, might be essential for the conservation of P. macrobullaris in the region.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Plecotus macrobullaris ( Kuzjakin, 1965 )
| Alberdi, Antton & Aizpurua, Ostaizka 2018 |
Plecotus alpinus
| Kiefer and Veith 2002: 8 |
Plecotus microdontus
| Spitzenberger 2002: 5 |
Plecotus auritus macrobullaris
| Kuzjakin 1965: 76 |