
Dystomanema Bezerra, Pape, Hauquier, Ingels & Vanreusel
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3692.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:F9EC43F2-F82E-4E8C-A735-ECA4DFF7CA00 |
|
DOI |
https://doi.org/10.5281/zenodo.6154051 |
|
persistent identifier |
https://treatment.plazi.org/id/707787ED-FFE7-2F1B-FF73-FF3CFEBCFDD2 |
|
treatment provided by |
Plazi |
|
scientific name |
Dystomanema Bezerra, Pape, Hauquier, Ingels & Vanreusel |
| status |
|
Genus Dystomanema Bezerra, Pape, Hauquier, Ingels & Vanreusel gen. nov.
Diagnosis. Neotonchinae . Body long, cylindrical, tapering anteriorly. Cuticle transversally striated with fine punctation, more evident in the tail region. No lateral differentiation. Few somatic setae with no fixed distribution pattern. Anterior sensilla arranged in three circles 6+6+4, first circle papilliform, outer circles setiform. Dorsosublateral setae just posterior to the third circle of sensilla at the level of the amphid. Spiral amphid smaller in females than in males. Oral opening ventrally displaced. Buccal cavity wide, deep, funnel shaped. Gymnostom followed by well-cuticularized pharyngostom with expanded conical posterior part. Three small teeth, one dorsal tooth and two smaller subventral teeth, all situated in the pharyngostom. Pharynx cylindrical with a posterior muscular bulb. Male reproductive system diorchic, spicules of equal size, one ventral precloacal spine and conspicuous cup-shaped precloacal supplements. Female reproductive system didelphic, ovaries antidromously reflexed. Tail conical.
Type species. Dystomanema cadizensis sp. nov.; present paper. One other species in genus: Dystomanema brandtae sp. nov.; present paper
Differential diagnosis. Dystomanema gen. nov. resembles Comesa Gerlach, 1956 and Neothonchus Cobb, 1933 in terms of the body shape, cuticle pattern and shape of supplements, but differentiates from both in having much smaller dorsal tooth.
In the subfamily Ethmolaiminae , Ethmolaimus de Man, 1880 (syn. Trichethmolaimus, Jensen 1994 ) and Paraethmolaimus Jensen, 1994 has long mouth and massive teeth; in the subfamily Neotonchinae , Comesa Gerlach, 1956 has a distinct dorsal tooth and ventral ridges; Gomphionema Wieser & Hopper, 1966 has a massive dorsal tooth; Gomphionchus Platt, 1982 and Neothonchus Cobb, 1933 has a big dorsal tooth; Nannolaimus Cobb, 1920 has a small weakly cuticularized dorsal tooth while Filitonchoides Jensen, 1985 and Filitonchus Platt, 1982 has a poorly developed buccal cavity with no sign of a dorsal tooth. Comesa Gerlach, 1956 species with small teeth also have a developed dorsal tooth. Dystomanema gen. nov. has no developed dorsal tooth but three small teeth (one dorsal and two subventral) instead. These small teeth are bigger than the ridges mentioned for Comesa Gerlach, 1956 . Dystomanema gen. nov. might represent a transition between no teeth ( Filitonchoides Jensen, 1985 and Filitonchus Platt, 1982 ) and small teeth (some Comesa Gerlach, 1956 ) to big teeth in this subfamily.
Differs from all genera of the family by the ventrally displaced oral opening (visible at low magnification).
Subterminal oral opening is observed in other nematoda genera, such as Campylaimus Cobb, 1920 , Diplopeltula Gerlach, 1950 and Morlaixia Vincx and Gourbault 1988 . Just some Campylaimus (Cobb, 1920) and Morlaixia Vincx and Gourbault 1988 (genus also found in cold-seeps by Guilini et al., 2012) have a subventrally displaced oral opening. Diplopeltula Gerlach, 1950 has a dorsal oral opening. The ventral position of the oral opening, in some Campylaimus , was not accepted by Vincx and Gourbault, 1988 when they described the genus Morlaixia Vincx and Gourbault, 1988 but Villares et al. (2013) accept this ventral position and provide a key to species identification.
Etymology. The genus is named after the particular mouth displacement, a characteristic that is unique in the family.
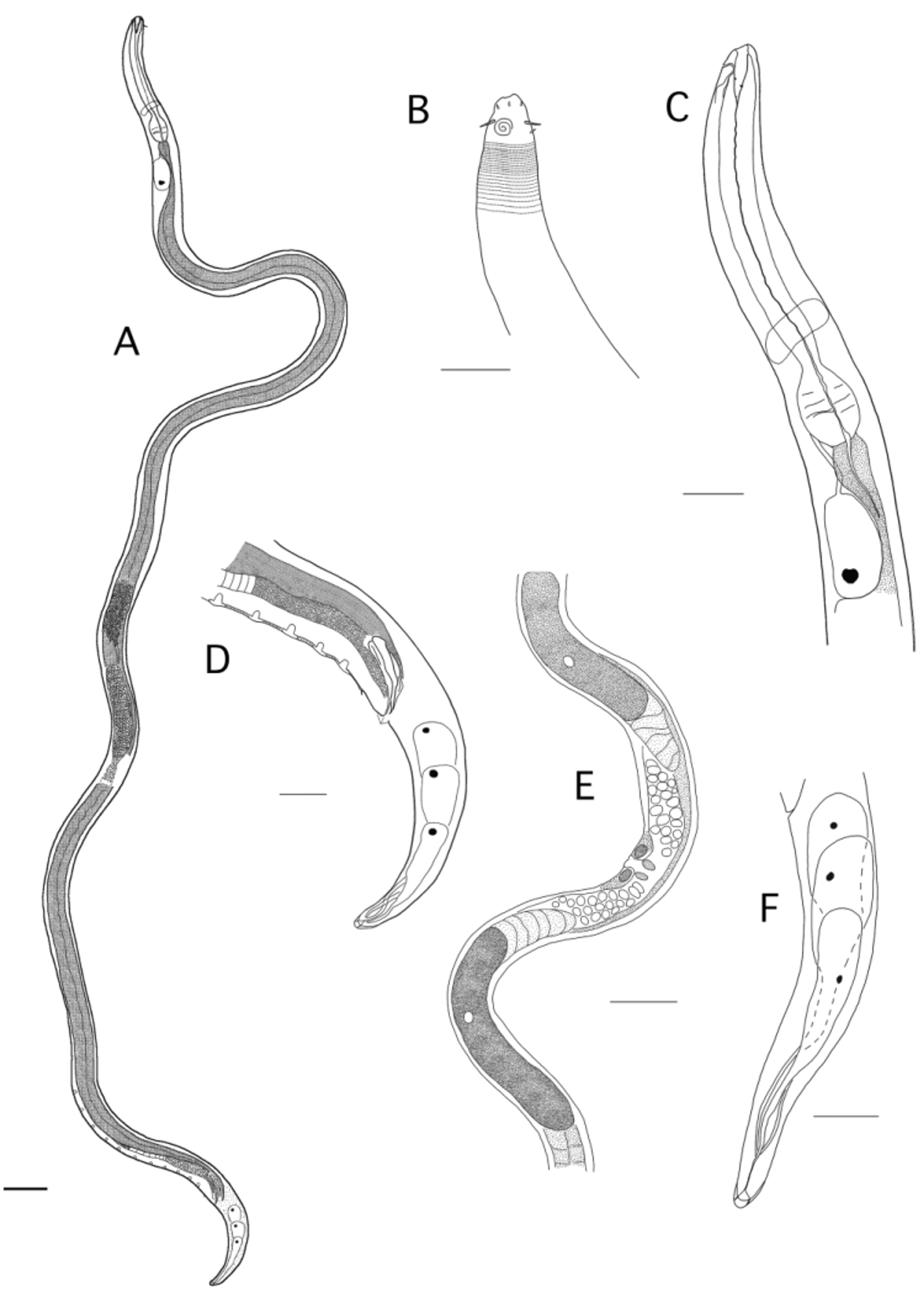
Dystomanema cadizensis gen. nov. sp. nov. ( Figs 4 View FIGURE 4 A–F, 5 A–H, 6 A–F; Table 1)
Material examined. Holotype male (Inventory No. UGMD 104262), three paratypes males (Inventory No. UGMD 104263, 104264, 104265), three paratypes females (Inventory No. UGMD 104266, 104267, 104268).
Type habitat and locality. Cold-seep sediments in the Northeast Atlantic, Gulf of Cádiz, Darwin mud volcano (35°23.51’ N 7°11.48’W).
Description. Males: Body slender, long, uniform in diameters tapering anteriorly and posteriorly. Fixation shape uncoiled. Cuticle transversally striated with fine punctation more evident on the tail region. No lateral differentiation. Few somatic setae with no fixed distribution pattern. Head conical, low lips. Inner labial sensilla papilliform, six outer-labial short setiform sensilla, four cephalic setae 5 µm or 35% of corresponding body diameter long. Short dorso-sublateral subcephalic setae present. Amphid spiral, ventrally to the centre, 3 turns. Mouth opening ventrally displaced. Buccal cavity wide, deep (18 µm), funnel shaped. Gymnostom followed by well-cuticularized pharyngostom with expanded conical posterior part. Sclerotized walls of pharyngostom with three small teeth: a dorsal tooth and two smaller subventral teeth. Pharynx cylindrical with a posterior muscular bulb. Nerve ring located slightly anterior to middle of pharynx. Secretory excretory pore located immediately after cephalic setae. Reproductive system with long vas deferens, clear patch of variable position from the level of the 4th to the 8th supplement from the cloaca. Outstretched testes anteriorly directed, anterior one to the right of intestine and posterior one, smaller, to the left. Spicules of equal size, about the same length as the cloacal diameter, slightly cephalated, bent about 1/3 of the distance from the distal end. Gubernaculum plate-like with a sclerotized rod-like internal structure at the distal half. Ten to twelve conspicuous cup-shaped precloacal supplements with external longitudinal articulated flange. Distance between the supplements variable among the males. Prominent ventral precloacal spine about 16 µm anterior to the cloacal opening. At the opening of the cloaca there is a slightly sclerotized triangular structure, which seems to be a cuticular projection. Tail conical plump, with bluntly rounded tip. Three caudal glands opening in a prominent, terminal blunt spinneret, half as wide as the tip of the tail.
Females: Females are similar to males in most respects, in general longer. Amphids do not show dimorphism in shape or number of turns but in females they occupy just 36% of the correspondent diameter, while in males this proportion is 45%. Vulva at 54% of the body; two perivulvar glands present, opening of perivulvar gland cells not clear; vagina transverse; ovejector sclerotized; vagina vera separated from vagina uterine by sphincter; uteri paired opposed; two cells (glands?) at the junction of vagina and uteri visible; spermatheca occupying about one third of each uterus; oviduct indistinct; ovaries antidromously reflexed, flexion at the junction of ovary and uterus. Anterior ovary to the left side of the intestine, posterior ovary to the right. Female tail conical, longer than in males. Terminal spinneret with same morphology than in males but slightly elongated.
Juveniles: similar to adults in most morphological aspects.
Differential diagnosis. Dystomanema cadizensis gen. nov. sp. nov. is characterized by its body size, number of supplements, both testes directed anteriorly and has the diagnostic features of the genus. The condition of both testes directed anteriorly was unknown within the family until now. Size of testes changes with male development but the posterior one always remains smaller than the anterior one. It is very close to Dystomanema brandtae gen. nov. sp. nov. but differs by the body size. Males about 70% smaller than males of D. brandtae gen. nov. sp. nov. Among females this difference is reduced to about 40%. The amount of precloacal supplements is also smaller: D. cadizensis gen. nov. sp. nov. has 10 to 12 cup-shape supplements while D. brandtae gen. nov. sp. nov. has 20.
Etymology. The species is named after the sampling location – Gulf of Cádiz.
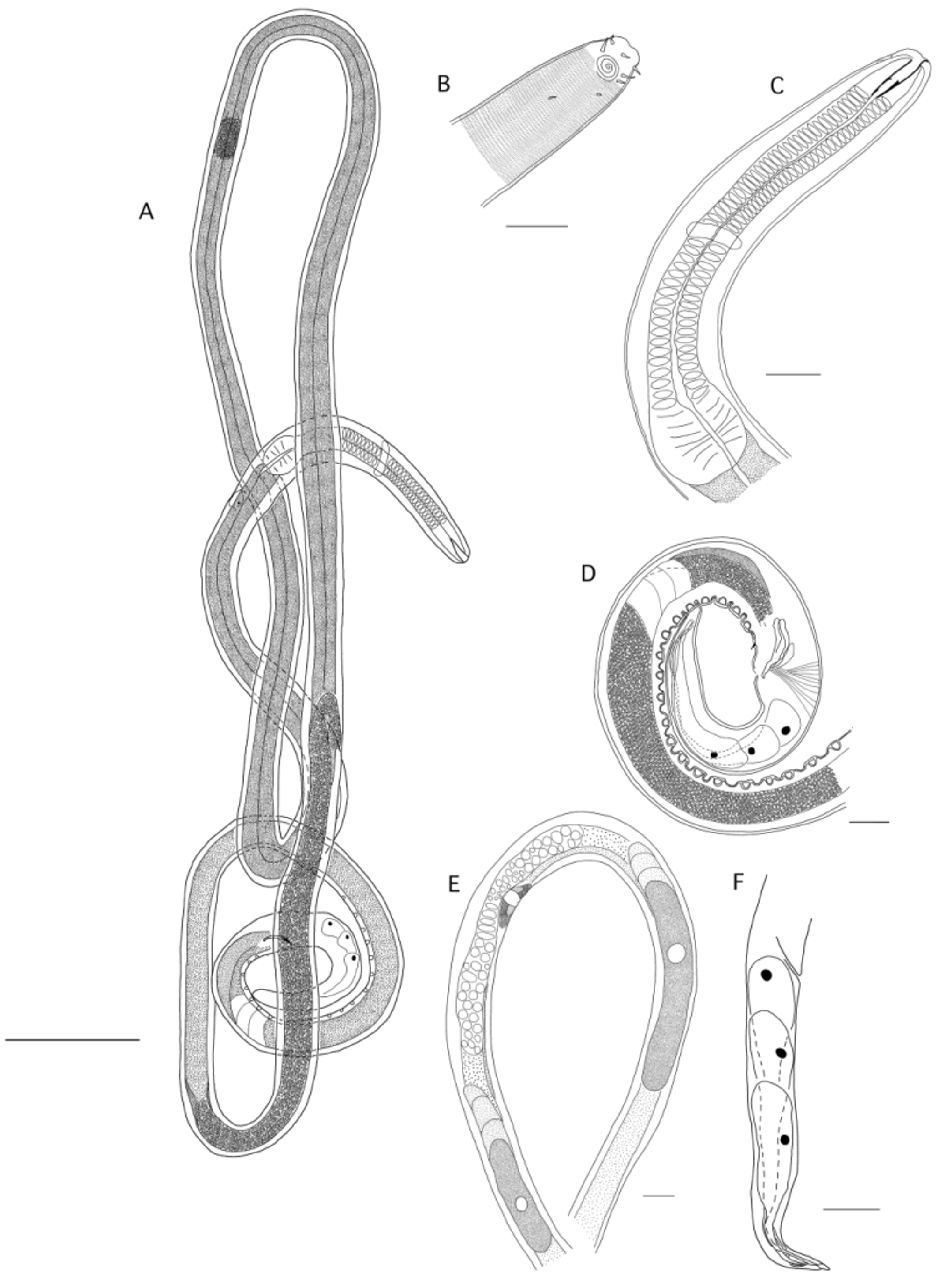
Dystomanema brandtae gen. nov. sp. nov. ( Figs. 7 View FIGURE 7 A–F, 8 A–F, 9 A–F, Table 1)
Material examined. Two males and 3 females. Holotype male (Inventory No. UGMD 104269), paratype male (Inventory No. UGMD 104270), three paratypes females (Inventory No. UGMD 104271, 104272, 104273)
Type habitat and locality. Remnant cold-seep sediments in the Larsen B area off the Eastern Antarctic Peninsula (65°26.07’– 65°26.10’S; 61°26.48’– 61°26.53’W)
Description. Males: Body slender, long, uniform in diameter, tapering anteriorly and posteriorly. Fixation shape mostly coiled. Cuticle transversally striated with fine punctation more evident on the tail region. No lateral differentiation. Few somatic setae with no fixed distribution pattern. Head conical, low lips. Inner labial sensilla papilliform, six outer-labial short setiform sensilla, four cephalic setae 4 µm or 25% of corresponding body diameter long. Short dorso-sublateral subcephalic setae present. Amphid spiral 3+ turns. Oral opening ventrally displaced. Buccal cavity wide, deep (21 µm), funnel shaped. Gymnostom followed by well-cuticularized pharyngostom with expanded conical posterior part. Sclerotized walls of pharyngostom with three small teeth, a dorsal tooth and two smaller subventral teeth. Pharynx cylindrical with a posterior muscular bulb. Nerve ring located about 20% of pharynx length from anterior end. Secretory excretory pore located immediately after cephalic setae. Reproductive system with long vas deferens, with clear patch from the level of the 7th to the 11th and 5th to the 10th supplement from the cloaca. Anterior testis outstretched and anteriorly directed, posterior one smaller and reflexed. Anterior testis to the right of intestine and posterior one to the left. Spicules of equal size, about the same length as the cloacal diameter, slightly cephalated, bent about 1/3 of the distance from the distal end. Gubernaculum plate-like with a sclerotized rod-like internal structure at the distal half. Twenty conspicuous cup-shaped precloacal supplements with external longitudinal articulated flange. Prominent ventral precloacal spine anterior to the cloacal opening. Tail conical plump, with bluntly rounded tip. Three caudal glands opening in a terminal blunt spinneret.
Females: Females are similar to males in most respects, in general longer. Amphids do not show dimorphism in shape or number of turns but in females amphids occupy just 40% of the correspondent diameter, while in males this proportion is 50%. Vulva at 53% of the body; four perivulvar gland cells present; vagina transverse; ovejector without special sclerotization; vagina vera inconspicuous, no sphincter present; uteri paired opposed; spermatheca developed as outpocketing of each uterus; oviduct indistinct; ovaries antidromously reflexed, flexion at the junction of ovary and uterus; ovary position in relation to the intestine not clear. Female tail conical, longer than in males. Spinneret terminal, elongated.
Juveniles: Similar to adults in most morphological aspects.
Differential diagnosis. Dystomanema brandtae gen. nov. sp. nov. is characterized by its body size, number of supplements, posterior testis reflexed and has the diagnostic features of the genus. The posterior testis is smaller than the anterior one. The condition of reflexed posterior testis has not been mentioned for the family until now. However, the drawing of Filitonchus ewensis description (Platt, 1982), shows a similar situation as observed in D. brandtae gen. nov. sp. nov. but he does not mention it and just describes it as “two opposed testes; anterior right, posterior left to the gut”. In addition, Lorenzen (1994) suggested that the posterior testis of Ethmolaimus (Family Ethmolaimidae , subfamily Ethmolaiminae ) is reflexed by describing the position of the testis structure from his own observations: “the posterior one (testis) proceeds anteriorly for a bit and then folds over to posterior”, although he doesn’t use the term ‘reflexed’. These observations suggest that the occurrence of a reflexed posterior testis has been overlooked in the past, and may support the classification of Dystomanema gen. nov. within the family Ethmolaimidae .
Dystomanema brandtae gen. nov. sp. nov. is very close to D. cadizensis gen. nov. sp. nov. but D. brandtae gen. nov. sp. nov. is about 70% longer, this difference is reduced to 40% when females of both species are compared. Unlike D. cadizensis gen. nov. sp. nov., the fixation shape of D.brandtae gen. nov. sp. nov. is mostly coiled. The punctation of D. brandtae gen. nov. sp. nov. remains fine but is more evident than in D. cadizensis gen. nov. sp. nov. Both species have very small teeth which can easily be overlooked, but in D. brandtae gen. nov. sp. nov. they are even smaller. D. brandtae gen. nov. sp. nov. has 20 cup-shaped supplements while D. cadizensis gen. nov. sp. nov. has 10 to 12.
The clear patch of the vas deferens mentioned in the revision of Ethmolaimidae (Platt, 1982) is also recognized in the two new species of the new genus. A similar structure has been described by Gourbault and Vincx (1988) for Calomicrolaimus compridus Gerlach, 1956 , although they mention “there is a clear patch because of the presence of only very fine granules in that region (cf. some Ethmolaimidae, Platt 1982 )”. In Dystomanema gen. nov. we didn’t find granules in this region. Muthumbi et al. (1997) also mention a clear patch for Sabatieria pisinna but do not explain if, like Gourbault and Vincx (1988), it is a region of very fine granules. The drawing of S. pisinna in the article doesn’t show the patch. Since the description of the patches in those other families does not match with the structures we identified in Dystomanema gen. nov., we are not able to affirm that the mentioned authors described the same type of structure.
Etymology. The species is named in honour of Prof. Angelika Brandt, for her valuable contributions to Antarctic research.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.