
Capros arambourgi, Baciu & Bannikov & Santini, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.5374196 |
|
persistent identifier |
https://treatment.plazi.org/id/720BF774-FFD6-6B1E-FF38-F9CD8E6ED5D7 |
|
treatment provided by |
Marcus |
|
scientific name |
Capros arambourgi |
| status |
sp. nov. |
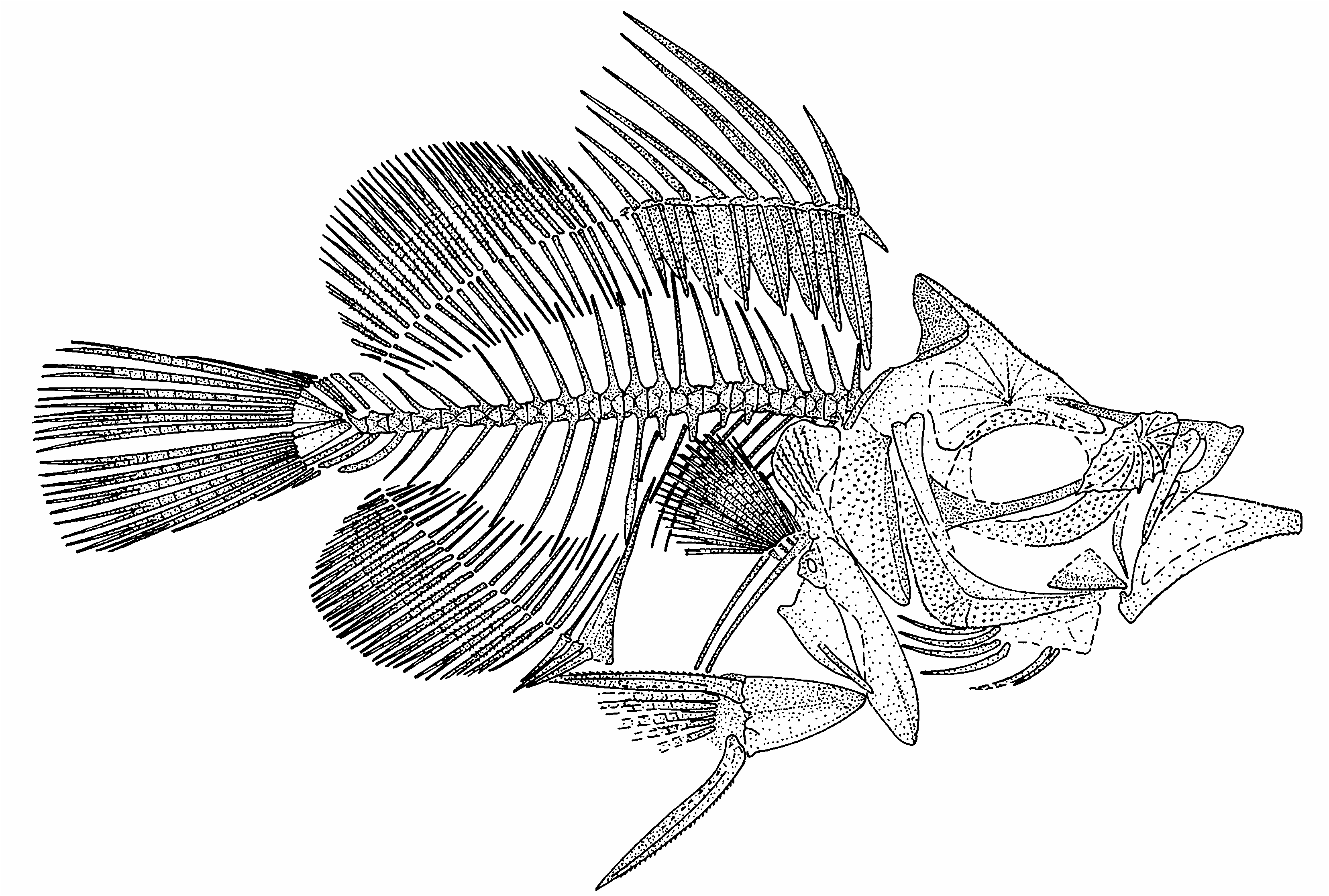
Capros arambourgi n. sp. ( Figs 1-3 View FIG View FIG View FIG )
Capros aper View in CoL – Arambourg 1921: 1243; 1927: 148, pl. XXVI, figs 2-4, pl. XXVII, fig. 1, text-figs 33, 34. — D’Erasmo 1930: III, figs 1, 2. — Bradley & Landini 1984: 30, pl. III, figs 4-6 (nec Capros aper (Linnaeus, 1758)) View in CoL .
Capros steindachneri – Arambourg 1927: 152, pl. XXVII, figs 2, 3, text-fig. 35 (nec Capros steindachneri (Gorjanović- Kramberger, 1882)).
HOLOTYPE. — MNHN ORA 85 D-G, part and counterpart, complete skeleton, 77 mm SL; upper Miocene , Messinian of Raz-el-Aïn (5.6-5 Ma), Oran, Algeria.
ETYMOLOGY. — The species is named in honour of Dr. Camille Arambourg, pre-eminent French paleontologist, in recognition of his contribution to the study of the Messinian fish fauna of Oran.
REFERRED SPECIMENS. — In the MNHN collection: in part and counterpart: ORA 17 D-G, ORA 98 D-G, ORA 733 D-G, ORA 741 D-G, ORA 743 D-G, ORA 747 D-G, ORA 749 D-G, ORA 1227 D-G and ORA 729/742 ; in single plate: ORA 16, ORA 86, ORA 90, ORA 103, ORA 395, ORA 563-566, ORA 730, ORA 731, ORA 736, ORA 739, ORA 740, ORA 744, ORA 746, ORA 747 D, ORA 748, ORA 752, ORA 1351; ORA 751 (two specimens in a single slab), skeletons and fragments, 46 to c. 90 mm SL, all from the type locality ; 2 specimens described by Arambourg (1927) as Capros steindachneri , ORA 15 (single plate) and ORA 396/1857 (part and counterpart), from the upper Miocene , Messinian of St-Denisdu-Sig, Oran, Algeria .
In the MSNTUP collection: 16 specimens from the upper Miocene, Messinian of Gabbro, Tuscany, Italy: MSNTUP I 10964 View Materials (3 skeletons on a single slab), I 10965 View Materials - I 10976 View Materials , I 11564, I 11588 and I 13994 ( I 10967 View Materials / I 10973 View Materials and I 10971 View Materials /I 11564 represent part and counterpart), their size ranges from 10 to 55 mm SL (most of the skeletons are more or less incomplete). According to d’Erasmo (1930), 10 specimens from the same locality of Gabbro are in the Museo Geologico di Bologna, and 2 specimens in the Museo Geologico di Firenze.
In the MPM collection: 1 uncatalogued specimen 46 mm SL, single plate, from the upper Miocene, Messinian of Mondaino, Emilia-Romagna, Italy.
TYPE LOCALITY AND HORIZON. — Most specimens examined originate from the type locality Raz-el-Aïn near Oran, NW Algeria. This locality is placed in the Djebel Murdjadjo carbonate platform in the Chelif Basin. The fish-bearing layers consist of light marls intercalated basally with limestone and are of Messinian (uppermost Miocene) age.
DIAGNOSIS. — Head length 2.2-2.6 in SL. Maximum body depth 1.8-2.0 in SL. The anterior angle of the supraoccipital crest extends to the vertical line through the posterior edge of the orbit or slightly anterior to it. There are 22 (10 + 12) vertebrae. The dorsal fin has nine spines and 20-21 soft rays; the third spine is the longest, 28-35% SL and 0.5-0.6 of maximum body depth. The sixth interneural space is vacant. The anal fin has three spines (the second spine being the longest) and 19- 20 soft rays. The soft dorsal- and anal-fin rays have spinules. The length of pelvic-fin spine is slightly greater than the distance between the posterior end of pelvis and the anterior edge of the first anal-fin pterygiophore.
DESCRIPTION
The body is deep, and has a short caudal peduncle. The caudal peduncle depth is about 0.10- 0.20 of the body depth. The head is relatively large, its length is about 0.6-0.8 of the body depth. The head length is contained 2.2- 2.6 times in SL. The maximum body depth is contained 1.8-2.0 times in SL. The dorsal and ventral profiles of the body are equally convex.
The head is deep, but its depth is not greater than its length. The eye is of moderate size, the horizontal diameter of the orbit being longer than the postorbital distance. The mouth is terminal, and presumably highly protractile. The lower jaw articulation is situated on the level vertical to the anterior edge of the orbit or slightly anterior to it. The infraorbital bones usually are not recognizable with confidence, but the lacrymal is often distinguishable. The lacrymal is thick, subrhomboid, and bears multiple spines and radiating ridges on its lateral surface, and there are strong serrations on the lower border.
Neurocranium
The neurocranium is deep, and the ethmoid region is moderately long. The supraoccipital crest is high, triangular, thorn-like, and pointed; its posterior edge is slightly concave, and the leading edge is thickened and lacks prominent ornamentation. The anterior angle of the supraoccipital crest extends to the level of the posterior edge of the orbit or slightly anterior to it. The frontal is highly ornamented over the orbit with radiating spinous crests. The parasphenoid is exposed in the lowermost part of the orbit; it is slender and almost straight, without a ventral flange.
New Caproidae (Teleostei) from the Miocene of Algeria
Oral jaws
The premaxilla has a long ascending process; the ascending process is evidently longer than the alveolar process. The alveolar process bears a well developed postmaxillary process. Small premaxillary teeth are distinguishable in some specimens. The maxilla is tapered and not expanded at its distal end.
The mandible is moderately long and low at the symphysis; its length being greater than its depth by about 2.0 times. The lower mandibular margin is slightly concave. There are small spines on the lower border of the dentary posterior to the symphysis. The retroarticular process of the angulo-articular has a posterior process directed along the lower border of the mandible. The lower jaw teeth are small.
Suspensorium
The hyomandibular shaft is inclined anteroventrally. The head of the hyomandibula is double in lateral view. The quadrate is triangular in shape,
with a small articular condyle. The ectopterygoid is curved, incompletely preserved.
Opercular region
The opercular region is narrow. The limits of the subopercle and interopercle are unclear. The opercle is vertically elongate; its lateral surface has a ridge which runs parallel to the anterior border of the bone. The opercle is indented posterodorsally, and it is ornamented laterally with radiating rows of spines. The preopercle is only slightly curved along its anterior edge, but the ventral margin of the dorsal limb (which is longer, albeit slightly) meets the posterior margin of the ventral limb almost at a right angle. The lateral surface of the preopercle is covered with spines. The peripheral border of the preopercle bears serrations and lacks a spine at the angle.
Hyoid and branchial arches
The epihyal is obscured in most specimens. The ceratohyal is partly recognizable in some specimens,
Baciu D.-S. et al.
and it has a distinctly concave ventral border. There are about six sabre-like branchiostegal rays. The pharyngeal teeth and branchial arch bones are not distinguishable in the material examined.
Axial skeleton
There are 10 abdominal and 12 caudal vertebrae. The first vertebra articulates closely with the skull (being “plastered” or tightly adherent to it). The vertebral column is almost straight, being slightly elevated anteriorly. The vertebral centra are almost square, being as long as high, and that of the first vertebra is fore-shortened. The neural spines of the abdominal vertebrae are fairly short, pointed, and either straight or slightly curved. The neural spines of the caudal vertebrae are slightly longer than those of the abdominal vertebrae, slender, and slightly curved; the spines become shorter posteriorly in the series. The haemal spine of the first caudal vertebra is somewhat stout, whereas the spines more posteriorly are slender. The neural and haemal spines of PU3 are slightly expanded distally. All the neural and haemal spines are inclined posteriorly, but those near the abdominal/caudal transition are close to vertical. Parapophyses are present on four of the posteriormost abdominal vertebrae. Pleural ribs are associated with the third to 10th abdominal vertebrae; they are very short and slender, increasing in length from the first to fifth; the penultimate rib is somewhat stouter than the preceding ones and the ultimate rib.
Pectoral fin and girdle
The posttemporal is not preserved. The supracleithrum is elongate and slender. The cleithrum is an elongate, robust and very slightly sigmoid bone situated just below the first vertebra. The ventral limb of the cleithrum is broad and spatulate; the dorsal limb tapers to a blunt point distally and it is expanded posteriorly as a broad plate ornamented laterally with fine serrate crests. The single postcleithrum is long and rod-like ventrally; the ventral end of the postcleithrum is close to the ventral profile of the body, and posterior to the pelvic bone. The coracoid is subtriangular and projects anteroventrally as a deep process which reaches the ventral limb of the cleithrum; the postcoracoid process seems short. There are four pectoral radials; the radials are slightly constricted in the middle and increase in length from above to below. The pectoral fin is inserted low on the flank, near the midpoint between the vertebral column and the ventral profile of the body. The pectoral fin probably is rounded distally; 15 pectoral-fin rays are recognizable in some specimens.
Pelvic fin and girdle
The pelvic bone is relatively long and wedge-like; it is oriented posteroventrally from its attachment to the cleithra. The posterior process of the pelvis is short. The pelvic fin is moderately long, with a strong spine and about five branched rays. The pelvic-fin spine and soft rays bear small spinules. The pelvic-fin origin is situated behind the pectoral-fin base. The pelvic-fin spine length slightly exceeds the distance between the posterior end of the pelvis and the anterior edge of the first analfin pterygiophore.
Dorsal fin
The dorsal fin is continuous, with a slight indentation between the spiny and soft portions. There are nine dorsal-fin spines; the first spine is the shortest. The first two spines are supernumerary on the first dorsal-fin pterygiophore. The third dorsal-fin spine is the strongest and longest; the succeeding spines gradually decrease in length posteriorly in the series. The third spine is 1.2 times longer than the fourth spine, and the fourth spine is somewhat longer than the pelvicfin spine. The second spine is less than one-half the length of the fourth spine. The six anteriormost dorsal-fin spines bear longitudinal striations. There are 20 or 21 soft segmented rays in the dorsal fin. The length of the base of the soft portion of the dorsal fin is slightly longer than the length of the base of the spiny portion. There are spinules on the soft rays. The soft dorsal-fin rays are more closely spaced than the dorsal-fin spines.
The pterygiophores of the spiny dorsal fin are large and sturdy, expanded anteroposteriorly, and each bears a longitudinal ridge. The first pterygiophore is the longest; its ventral shaft is situated in front of the first interneural space, probably being inserted between the halves of the bifid neural spine of the first vertebra. There is a point- ed procumbent spine directed anteroventrally from the upper part of the first dorsal-fin pterygiophore. The sixth interneural space is vacant. The ventral shafts of the dorsal-fin pterygiophores are inclined posteriorly. The pterygiophores of the soft dorsal fin are slender and decrease in length posteriorly in the series. Each interneural space between the 11th to 18th vertebrae contains two or three ventral shafts of the pterygiophores of the soft dorsal fin.
Anal fin
There are three spines and 19 or 20 soft segment- ed rays in the anal fin. The first two anal-fin spines are supernumerary on the first pterygiophore. The second spine is the longest, and the third spine is the shortest. The middle anal-fin soft rays appear to be the longest. The anal-fin soft rays bear spinules.
The first anal-fin pterygiophore is long and stout; its dorsal shaft is slightly inclined, being situated along the anterior edge of the haemal spine of the first caudal vertebra. The ventral end of this pterygiophore is slightly expanded anteriorly. All the other anal-fin pterygiophores are slender, most of them are almost as equally long as opposite soft dorsal-fin pterygiophores. The anal-fin pterygiophores are inclined posteriorly; their lengths gradually decrease posteriorly in the series. The interhaemal spaces above the anal fin have the dorsal shafts of two to four pterygiophores present.
Caudal fin and skeleton
The hypurals and parhypural are autogenous. The short neural spine of PU2 forms a low triangular crest. There are three epurals; uroneurals are not visible in the material examined. The centrum of PU2 is only slightly foreshortened; its haemal spine is autogenous.
The caudal fin is long and rounded; it has 14 principal rays (I,6 + 6,I), and two procurrent rays both above and below.
Squamation
The body is completely covered with scales which bear numerous (up to 20) needle-like laterally projecting spines on their surfaces. The scales are relatively small and either round or oval, with even margins. The lateral line is not evident in most of the specimens; a few specimens show the arched anterior part of the lateral line.
Measurements (see Table 1)
TABLE 1. — Measurements taken from six specimens of Capros arambourgi n. sp. of 46-85 mm standard body length (SL).
% ofSL Head length from tip of snout to posterior border of opercle 38-45 Maximum body depth 50-58 Depth of caudal peduncle 9-12 Distance between origins of pelvic and anal fins 17-22 Length of first spine of dorsal fin 2.5-3.5 Length of longest spine (third) of dorsal fin 27-38 Length of base of spiny dorsal fin 22-26 Length of base of soft dorsal fin 23-27 Length of longest spine (second) of anal fin 7-14 Length of base of anal fin 30-33 Length of spine of pelvic fin 22-28 Length of longest ray of caudal fin 28-30 Length of lower jaw 15-19
REMARKS
Our investigation of the MNHN Messinian Capros materials from Oran ( Algeria) revealed several incorrect interpretations of characters made by Arambourg (1927) in his descriptions of C. aper . For example, the vertebral formula of the Messinian boarfish is 10 + 12 rather than 9(10) + 13 as indicated by Arambourg (1927: 151). Also, none of the specimens available have more than 21 soft rays in both the dorsal and anal fins (Arambourg indicated 23 soft dorsal- and analfin rays).
I n a d d i t i o n t o C. a p e r, A r a m b o u r g (1 9 2 7) described from the Messinian of Oran the species C. steindachneri (Kramberger, 1882) based on two specimens from the site Saint-Denis-du-Sig. We refer both specimens to C. arambourgi n. sp. because the supposedly distinctive characters of “ C. steindachneri ” either lie within the range recorded in the new species (body depth, relative lengths of the third dorsal-fin spine and pelvic- and anal-fin spines) or can be explained by a slight distortion of the neurocranium (the “profile fronto-facial” relatively more abruptly oblique). The species name steindachneri should be regarded as a synonym of the Sarmatian species Proantigonia radobojana Kramberger, 1882 from Croatia ( Baciu et al. 2005).


| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Capros arambourgi
| Baciu, Dorin-Sorin, Bannikov, Alexandre F. & Santini, Francesco 2005 |
Capros steindachneri
| ARAMBOURG C. 1927: 152 |
| BRADLEY F. & LANDINI W. 1984: 30 |
| ARAMBOURG C. 1927: 148 |
| ARAMBOURG C. 1921: 1243 |