
Xyela graeca J.P.E.F. Stein, 1876
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3629.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FF47F026-9CB6-4390-B900-130A3DF2B33B |
|
DOI |
https://doi.org/10.5281/zenodo.6412594 |
|
persistent identifier |
https://treatment.plazi.org/id/72655041-410A-E90A-6BCD-D161FBF120C1 |
|
treatment provided by |
Felipe |
|
scientific name |
Xyela graeca J.P.E.F. Stein, 1876 |
| status |
|
Xyela graeca J.P.E.F. Stein, 1876
Xyela graeca J.P.E.F. Stein, 1876: 57–58 , ♀, type locality: Greece, Nomos Ioanina, Konitsa S 6 km; Konow 1897: 58 (junior synonym of julii); Benson 1938: 33, 35 (removed from synonymy).
Pinicola graeca : André 1881: 468 (combination with Pinicola).
Xyela nigrae Rasnitsyn, 1965: 519 , ♀, type locality: Ukraine, Transcarpathia, Tur’i-Remety near Perechin; Rasnitsyn 1971: 193 (junior synonym of graeca).
Description. Female. Color. Head yellow with black and brown pattern: black stripes along frontal furrows often fading and at most 2 times wider than ocellar diameter, meeting black ocellar and postocellar area, black longitudinal medial spot of frons always present, weak in pale specimens, rarely fusing with frontal stripes, kidneyshaped spots on vertex separate from black postocellar area ( Fig. 51–52 View FIGURES 42–65 ). Antennae brown or pale brown, scape sometimes yellow. Thorax dorsally brown with rich yellow pattern on pronotum, mesonotal lobes and mesoscutellum, tegulae yellow and sometimes brown in middle, mesepisternum largely pale yellow. Abdominal terga brown, lateral parts of terga 7, 8 and 9+10 yellow or pale brown, valvifer 2 completely yellow or pale brown, membrane between valvifer 2 and valvula 3 white, valvula 3 varying from yellow to pale brown ( Fig. 107 View FIGURES 105–120 ). Legs pale brown, posterior coxae pale brown or yellow and more or less infuscate from base. Wing membrane clear, venation and pterostigma pale.
Morphology. Fore wing 2.6–3.9 mm long, 1.90–2.35(–?2.55) times longer than ovipositor sheath, vein Rs+M 130–280 µm long, 2r-m meeting Rs proximal to furcation of Rs1 and Rs2. Synantennomere 3 510–710 µm long, antennomere 4 130–160 µm long and 4.5–5.5 times longer than wide distally. Article 3 of maxillary palp 410–480 µm long, 1.65–1.85 times longer than scape and wider than synantennomere 3. OOL: POL = 1.60–2.00: 1. Ovipositor sheath (1.35–) 1.60–1.80 mm long, valvula 3 (?1.60–)1.70–2.00 times longer than valvifer 2 and (?4.5–)4.8–5.8 times longer than wide at base ( Fig. 107 View FIGURES 105–120 ). Valvula 3 of ovipositor compressed, pale membranous area about as long as basal width of valvula 3, dorsal edge of valvula 3 sloping down to round tip, distally with sensilla field exposed and directed caudally, bearing 3 setae. Ovipositor straight and compressed. Valvula 1 with aulax terminating distally, ventral edge sloping up to tip, with ca 13–15 closely spaced oblique annuli in distal quarter, without serrulae, olistether with 4–5 setae. Left and right valvulae 2 fused along dorsal edge in basal half. Valvula 2 with smooth dorsal margin, tapering in distal half, pale and evenly sclerotized, in distal 0.4 with single scattered sensilla campaniformia, in distal 0.1 with 3–5 oblique annuli. Posterior tibia 0.80–1.05 mm long, claws without subapical tooth.
Male. Color. Similar to female, but often generally paler (see Fig. 53 View FIGURES 42–65 for color pattern of head). Hypopygium yellow, often also preapical sterna largely yellow.
Morphology. Fore wing 2.9–3.5 mm long, Rs+M 160–250 µm long, 2r-m meeting Rs (130–)180–280 µm proximal to furcation of Rs1 and Rs2. Synantennomere 3 510–700 µm long, antennomere 4 140–200 µm long and 5.5–7.0 times longer than wide distally. Article 3 of maxillary palp 340–430 µm long, 1.35–1.55 times longer than scape and wider than synantennomere 3. OOL: POL = (1.40–)1.60–2.00(–2.20): 1. Longitudinal apodeme of basiparamere curved, basal portion in lateral position, harpe about as long as wide in lateral view. Lower ergot on valvular stalk absent. Valviceps (1.70–)1.80–2.00 times longer than wide on medial lobe, lateral lamella distinct and oblique, proximal lobe of penis valve at proximal end round, 0.20–0.30 times as long as valviceps and 0.70–0.80 times as high as medial lobe, excision on lower edge 0.16–0.22 as deep as width of medial lobe, valviceps on medial lobe 1.35–1.55 times wider than on distal lobe, 2 distal flagella present, tip of longer flagellum reaching (0.90–)0.95–1.05 width of distal lobe ( Fig. 142 View FIGURES 136–145 ). Valviceps with median longitudinal sclerotization present, medial lobe almost symmetrical and broad, with 5–10 cone-like sensilla along upper edge and scattered on lateral surface, upper edge between medial and distal lobe with 6–9 setae. Posterior tibia 0.70–0.85 mm long, claws without subapical tooth.
Barcodes. GUID ABU8786 View Materials (2♀ 1♂)
Type material. Xyela graeca . Neotype ♂ (here designated): “ Greece, Nomos Ioanina, Konitsa S 6 km, 650 m NN [altitude above sea level], 40°06’N 20°46E, leg. Blank & Kutzscher c. [collected] 4.5.1999 em. [emerged] 21.3.2000 [rearing number] Z37/99, larva ex Pinus nigra [i.e., Pinus nigra pallasiana ]”; “ Xyela graeca J.P.E.F. Stein, 1876 det. S. M. Blank 2000”; [rot:] “Neotypus ♂ Xyela graeca J.P.E.F. Stein, 1876 des. S. M. Blank 2001”. Genitalia stored inside small vial and kept on pin of type specimen. DEI. GoogleMaps
Xyela nigrae . Holotype ♀: [ Cyrillic letters:] “ Karpati , Tur’i-Remety bliz Perechina [= Tur’i-Remety near Perechin ], 15.V.65, A. Rasnitsyn ”; [rot:] “ Holotypus Xyela nigrae ♀ A. Rasnitsyn ”. In good condition. ZMUM . Paratype: 1♀, ZMUM .
Host plant. Pinus brutia Ten. ( Schedl 1981) , Ο Pinus nigra ssp. laricio Poir , P. nigra ssp. mauretanica Maire & Peyerimhoff ( Berland 1937, 1943), ● Pinus nigra ssp. nigra Arn. (43♀ 21♂ from 6 reared samples), ● P. nigra ssp. pallasiana Lamb. (78♀ 28♂ from 9 reared samples), Ο P. nigra ssp. salzmannii Dunal , P. sylvestris L. (2♂ from 1 reared sample).
Biology. In a series reared from two almost syntopic samples of Pinus nigra pallasiana taken in southern Turkey X. menelaus hatched significantly earlier than X. graeca (females: n[ graeca ] = 10, n[ menelaus ] = 14, U = 9, α <0.001; males: n[ graeca ] = 5, n[ menelaus ] = 11, U = 7, α <0.05; U-test). The difference was on average 6 days. The few X. curva from this sample hatched even ca 18 days earlier than X. graeca . This corresponds well with Pschorn-Walcher & Altenhofer’s (2000) observation, that Austrian X. graeca emerge in April, 1 or 2 weeks later than X. curva . In fact, Pschorn-Walcher & Altenhofer (2000) had a mixture of X. graeca and X. menelaus on hand.
For additional phenological data see Blank (2002). Specimens collected in February 1937 in Algeria ( Berland 1937 recording X. julii from Tikdja in the Djurjura) were identified as X. graeca by Blank (2002) but they might belong to a species associated with Pinus halepensis .
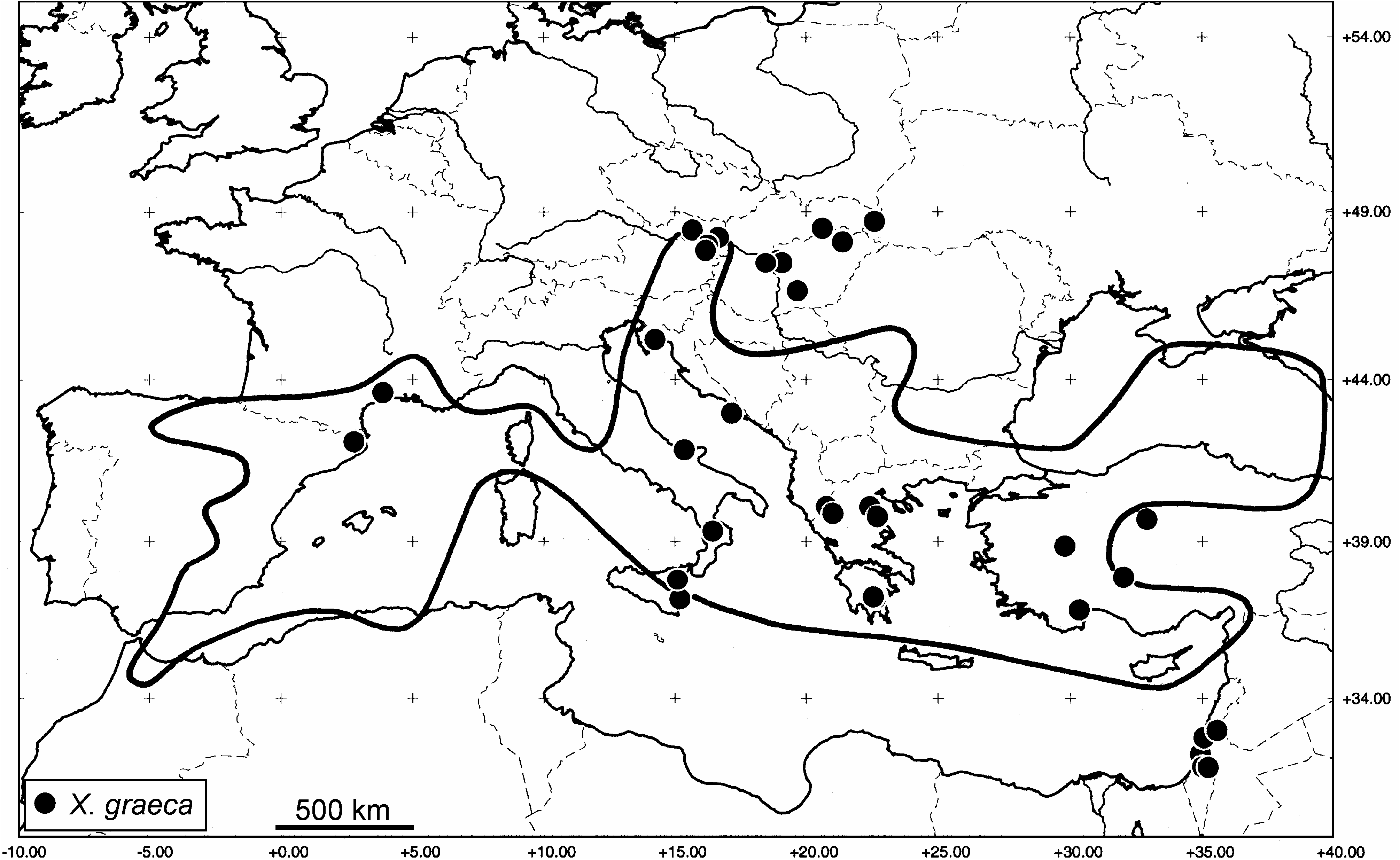
Geographic distribution. Austria, Croatia, France, Greece, Hungary, Israel, Italy, Spain, Turkey, Ukraine ( Fig. 15 View FIGURE 15 ). Additionally reported from Bulgaria ( Vasilev 1987), the Slovakian Republic ( Roller 1999) and Cyprus (Schedl 2002 as X. “ cf. graeca ”). These records are likely to be correct but should be re-examined, because they could relate to X. graeca or to X. menelaus . The record for Corsica by Liston & Späth (2005) refers to X. menelaus .
Remarks. Among the pale representatives of the Xyela julii group with a comparatively short ovipositor two very similar forms occur in the West Palearctic, which both feed on Pinus nigra . Corresponding males have an apparent longitudinal sclerotization of the valviceps and a gradually ascending distal edge of the valviceps. The existence of two separate species is well founded upon the recognition of two different forms of penis valves. These species are here called X. graeca and X. menelaus , and in accordance with Benson (1960) the name menelaus is applied to the species with the shorter ovipositor (see below for discussion on nomenclature). The sexes of the two species were associated with the help of two reared series from Lower Austria and a reared series from southern Turkey. The first series from Hernstein mainly comprised of females with a comparatively long ovipositor and males with a shallowly excised valviceps bearing a narrowly rounded proximal lobe (= X. graeca ). The second series from Dürnstein contained mainly females with shorter ovipositor and males with deeper excised valviceps having the proximal side of the proximal lobe truncate (= X. menelaus ). This association of the sexes corresponds with the emergence phenology of specimens from the reared Turkish series (see above and Fig. 14 View FIGURE 14 ).
Unlike the males the discrimination of females of X. graeca and X. menelaus is often dubious. The relative length of fore wing: ovipositor sheath and valvifer 2: valvula 3, and the absolute ovipositor sheath length exhibit bimodal distributions. Since a wide overlap is considered, the characters in the key will separate some 80 % of the females.
In the barcoding analysis, three specimens of X. graeca were grouped in a common cluster with 0.93 % intraspecific variability. This cluster is well separated from the cluster comprising specimens identified as X. menelaus ( Fig. 23 View FIGURE 23 ). The closest neighbor of X. graeca is X. obscura at an interspecific distance of 6.13 %.
Xyela graeca was described from two females collected in Greece. Stein (1876) reported his own collection as their place of deposition. Today the collection of J.P.E.F. Stein is preserved at ZMHB ( Groll 2010), but no corresponding specimen could be found there. Already Benson (1938) reported that H. Bischoff, former curator of the Hymenoptera section of ZMHB, was not able to trace the X. graeca type material, “as it was never received there with the rest of the collection”. As types of other taxa described by Stein (1876) like Lyda maculipennis (= Kelidoptera m.; see Blank et al. 1998) and Pachycephus smyrnensis can still to be located in ZMHB, the syntypes of X. graeca are deemed to be lost.
Stein’s (1876) pale Xyela species can be assigned to the X. julii group because of its knife-like ovipositor sheath. Konow (1897) regarded it as a “schwächliches Exemplar” (feeblish specimen) of X. julii . Enslin (1918) cast doubt on the correctness of Konow’s opinion referring to Stein’s description of the comparatively short and yellow ovipositor sheath. Benson (1938) removed X. graeca from synonymy and applied the name to a Mediterranean species, which is distributed northwards to Austria. Already Berland (1943) considered Stein’s description not to allow the recognition of this species, and the assignment of the name to a particular species to remain uncertain as the type is lost. But he maintained the name for a Mediterranean species. Benson (1960) added a very similar species from Greece, X. menelaus , which was considered to differ from all previously known in the world by its very short ovipositor. He omitted to give a reason, why he believed it to be different from Stein’s X. graeca , which was also described due to its short ovipositor. Actually, Stein’s description suits both, X. graeca and X. menelaus , and both may be common on Greek stands of Pinus nigra . Interpreting only the original description it is impossible to decide which of them Stein had on hand, as he did not consider the exact proportion of the ovipositor sheath. The designation of a neotype is necessary to promote nomenclatural stability by assigning the name X. graeca to a distinct Mediterranean species of the X. julii group. Differing from the original type series a male specimen has been chosen as a consequence of the ambiguous identification of females (Art. 75.3.5., ICZN 1999). Like the original types it originates in Greece. It was reared from Pinus nigra pallasiana .
In a footnote Berland (1943) mentioned X. graeca collection material from the MNHN labeled as “ pinicola, n. sp.” by Abeille de Perrin. However, the name is not available as “ pinicola ”, because it refers to Pinicola which has been assessed as a valid generic name in former times (see Berland 1947, who refers to the same specimen citing it as “ Pinicola , n. sp. ” written with a capital initial letter).
Study of the X. nigrae holotype confirms the synonymy of this name with X. graeca , which has already been proposed by Rasnitsyn (1971). Both X. nigrae type specimens were reared from Pinus nigra .
Schedl (1981) reported dark “ X. graeca ” females collected from P. brutia near Thripi on the Greek island Crete, and accordingly he concluded P. brutia to be an additional host plant of X. graeca . Berland (1937 under the name X. julii, 1943 ) accounted P. nigra var. mauretanica as a host of X. graeca , but the presence of X. graeca in Morocco or Algeria has not been confirmed. Such material belongs to forms which are similar to X. altenhoferi ( Fig. 13 View FIGURE 13 ) and these host record should be deleted for X. graeca .
Single specimens of X. graeca occurred in a reared series of X. julii from P. sylvestris . The sample was taken from a mixed stand of P. sylvestris and P. nigra in Lower Austria. In northeastern Germany P. sylvestris was found to shed pollen about two weeks earlier than P. nigra , but after a cold spring the phenology of both pines may overlap providing females the opportunity to oviposit on an unusual pine species.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Xyela graeca J.P.E.F. Stein, 1876
| Blank, Stephan M., Shinohara, Akihiko & Altenhofer, Ewald 2013 |
Xyela graeca J.P.E.F. Stein, 1876: 57–58
| Benson, R. B. 1938: 33 |
| Konow, F. W. 1897: 58 |
| Stein, J. P. E. F. 1876: 58 |