
Peruanocotyle pelagica Ruiz-Escobar, Torres-Carrera, and Ramos-Sánchez, 2022
|
publication ID |
https://doi.org/ 10.1645/21-27 |
|
publication LSID |
lsid:zoobank.org:pub:8EB566BC-B325-4837-B752-5612DD4DA35D |
|
DOI |
https://doi.org/10.5281/zenodo.7753840 |
|
persistent identifier |
https://treatment.plazi.org/id/7301879F-FF84-1906-FCBE-FA92F62792A2 |
|
treatment provided by |
Felipe |
|
scientific name |
Peruanocotyle pelagica Ruiz-Escobar, Torres-Carrera, and Ramos-Sánchez |
| status |
sp. nov. |
Peruanocotyle pelagica Ruiz-Escobar, Torres-Carrera, and Ramos-Sánchez View in CoL View at ENA n. sp.
( Figs. 1–10 View Figures 1–3 View Figures 4–8 View Figures 9, 10 )
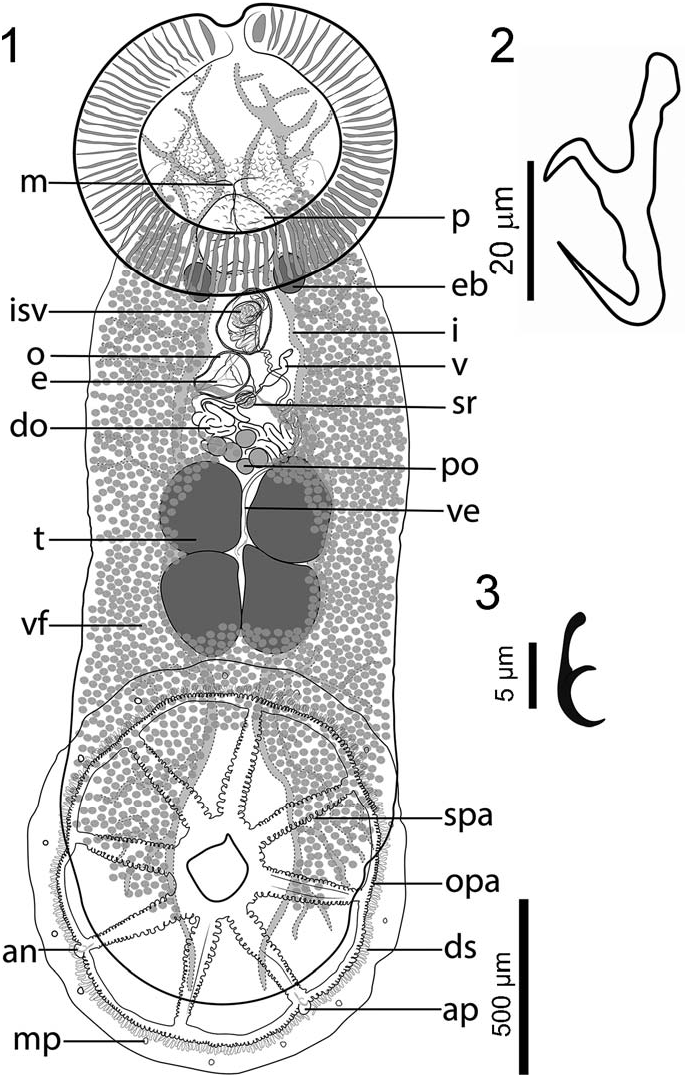
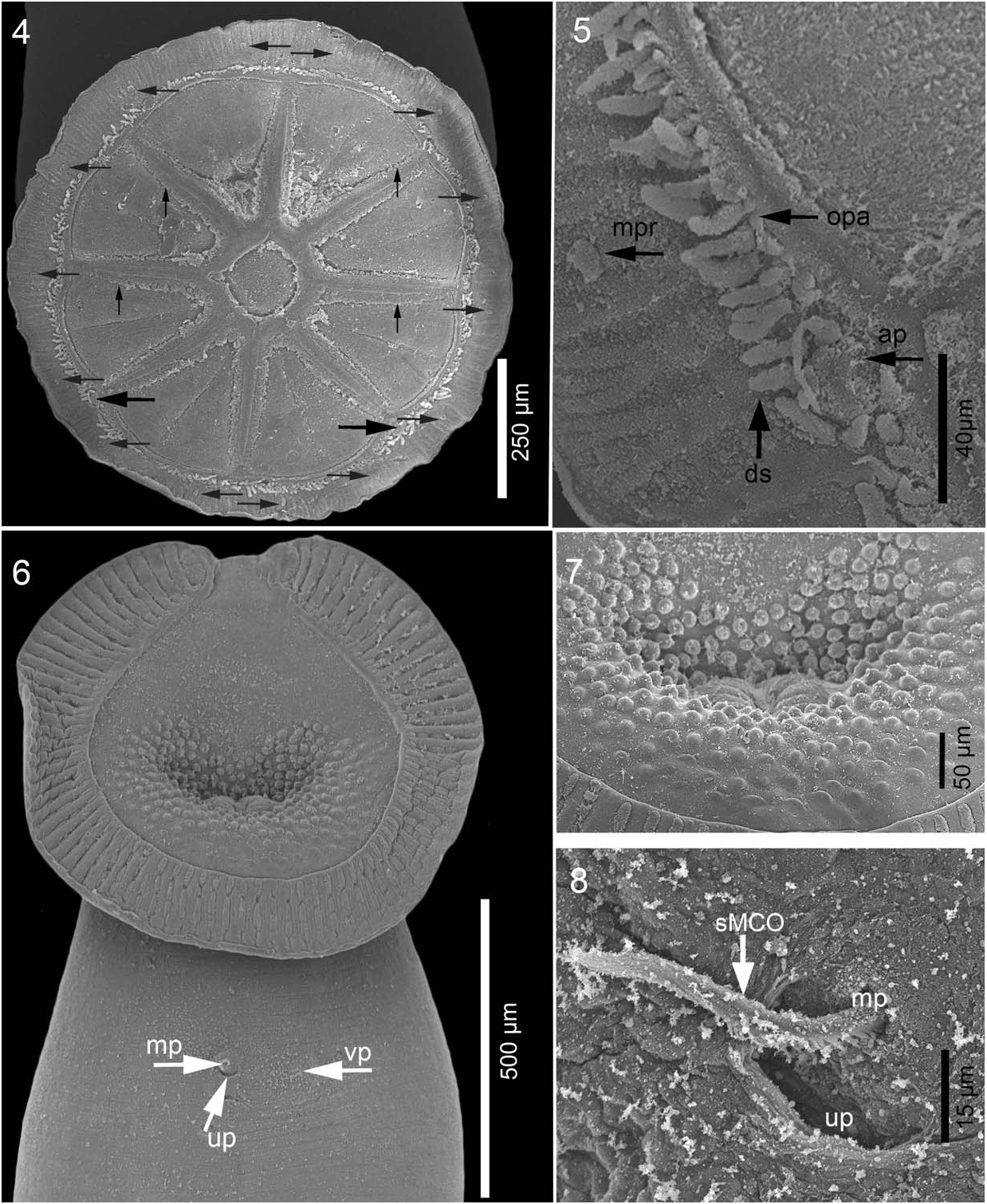
Description based on 16 specimens: Body whitish, translucent in vivo, elongate, including haptor 2,500 (1,500 –3,500, n ¼ 9) length, 790 (420–1,330, n ¼ 9) maximum width at posterior third of trunk, slightly constricted at level of testes ( Fig. 1 View Figures 1–3 ). Tegument smooth. Haptor wheel shaped, 880 (600–1,300, n ¼ 6) long by 854 (600– 1,260, n ¼ 6) wide, with 1 central loculus and 8 peripheral loculi. Marginal membrane (valve) irregular in width, 92 (67–124, n ¼ 4) ( Fig. 1 View Figures 1–3 ). A pair of small anchors, handle shorter than blade, rounded tip; slender guard curved towards the tip of blade, without accessory sclerotized piece ( Fig. 2 View Figures 1–3 ). Each anchor [44 (41– 52, n ¼ 6) long] embedded in processes, located at postero-lateral radial septa, slightly protruding into marginal membrane. Marginal membrane with 14 hooklets, 12 long, 1 or 2 per loculus as illustrated ( Fig. 3 View Figures 1–3 ), embedded in small processes, 10 wide ( Figs. 4, 5 View Figures 4–8 ). Radial and outer-ring septa, with numerous haptoral papillae. Marginal membrane with numerous dendritic structures, protruding into valve approximately 20 long by 5 wide ( Figs. 4, 5 View Figures 4–8 ). Anterior attachment organ well developed, ovoid, 728 (350– 1,130, n ¼ 7) long and 815 (470–1,350, n ¼ 7) wide, with 86 (81–92, n ¼ 3) oblique muscle grooves and antero-medial notch ( Fig. 6 View Figures 4–8 ). Anterior glands, ducts, and openings not observed. Mouth midventral, with around 297 dome-shape papillae surrounding oral opening, 18 (16–20, n ¼ 5) diameter ( Fig. 7 View Figures 4–8 ). Eyespots absent. Genital pores open adjacent, slightly to right of the body’s midline ( Figs. 6, 8 View Figures 4–8 ).
Pharynx semi-ovate 267 (187–340, n ¼ 6) long and 257 (155– 350, n ¼ 9) wide. Esophagus short, almost indistinct. Cecum bifurcation anterior to ejaculatory bulb. Intestinal ceca with two main lateral branches on each side of body. Each branch with abundant lateral diverticula, extends to posterior end of body proper. Pair of oval excretory bladders, 179 (110–250, n ¼ 6) long, 117 (60–160, n ¼ 6) wide, on each side of posterior end of pharynx ( Fig. 1 View Figures 1–3 ).
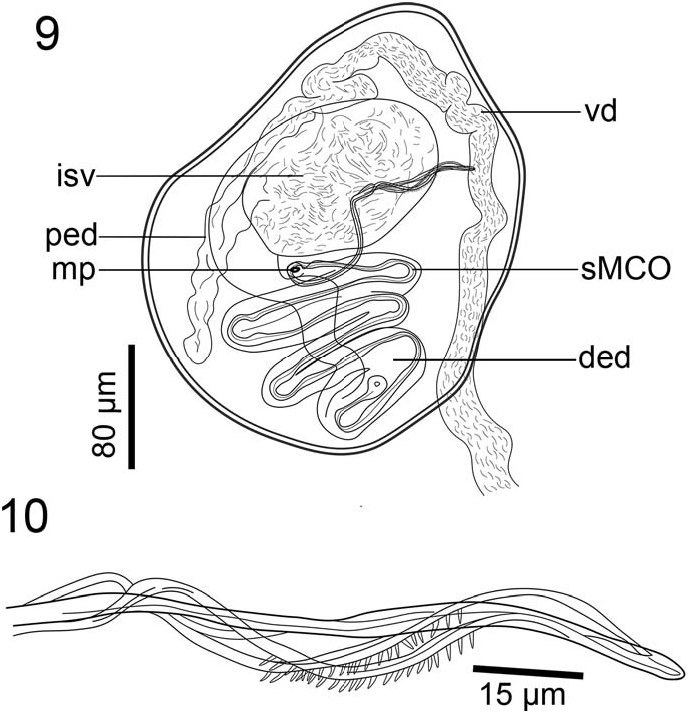
Four testes arranged in 2 pairs, oval to angular, equally sized and independent, 240 (112–430, n ¼ 13) long and 200 (120–340, n ¼ 13) wide, occupying intercecal space ( Fig. 1 View Figures 1–3 ). Vas deferens convoluted, running dorsal to vagina and entering anterior end of ejaculatory bulb; external seminal vesicle not observed. Seminal vesicle semi-globular, within ejaculatory bulb, 147 (143–151, n ¼ 4) long and 94 (92–97, n ¼ 4) wide. Ejaculatory bulb muscular, thickwalled, semi-oval, 273 (254–303, n ¼ 5) long and 191 (169–202, n ¼ 5) wide ( Figs. 1 View Figures 1–3 , 9 View Figures 9, 10 ). Ejaculatory duct sinuous, thick-walled, proximal and distal portions differentiated. Distal portion thicker at its origin, extending posteriorly from the internal seminal vesicle and slightly sinuous before forming the distal portion, which is convoluted and anteriorly directed ( Fig. 9 View Figures 9, 10 ). Male accessory glands not seen. Male copulatory organ (MCO) sclerotized, whip shaped, arising from distal end of ejaculatory bulb, inside of zig-zag portion of distal ejaculatory duct ( Fig. 9 View Figures 9, 10 ). MCO ending in 4 interlaced extensions, connected at end, two of them armed with 20 thin spines each, 2 long (2–3, n ¼ 2) ( Figs. 9, 10 View Figures 9, 10 ).
Ovary ( Fig. 1 View Figures 1–3 ) midventral with distinct elongated and coiled distal portion; proximal portion with 5 spherical lobes 60 (51–67, n ¼ 3) in diameter. Vagina single, opening ventrally on left side, at level of distal end of ejaculatory bulb. Vaginal pore unarmed. Non-sclerotized vaginal duct, undifferentiated, thick walled, Sshaped, narrows immediately before entering seminal receptacle; glandular cells not seen. Oötype ovoid, 122 (122–181, n ¼ 3) long and 96 (96–194, n ¼ 3) maximum wide, posterior to male and uterine pores. Seminal receptacle 91 (90–97, n ¼ 5) long and 71 (62–87, n ¼ 5) wide. Vitellarium extends from pharynx region to end of body proper, with abundant vitelline follicles, partially obscuring lateral branches of ceca; some follicles invade testicular field ( Fig. 1 View Figures 1–3 ). Egg, not measured, tetrahedral in shape.
Taxonomic summary
Type host: Rhinoptera steindachneri Evermann and Jenkins, 1891 ( Myliobatiformes : Rhinopteridae ), Pacific cownose ray.
Type locality: Off Salchi , Oaxaca, Mexico (15°40 ′ 51 ′′ N, 96°20 ′ 48 ′′ W), Tropical Eastern Pacific GoogleMaps .
Other localities: Off Ventanilla , Oaxaca (15°39 ′ 52 ′′ N, 96°34 ′ 31 ′′ W), Acapulco, Guerrero, (6°51 ′ N, 99°52 ′ W) GoogleMaps .
Type material: Holotype: CNHE 11461 ; paratypes: 5 specimens CNHE 11462 (off Salchi, Oaxaca, 14 December 2019); UMARPLAT 045 (8 specimens, off Ventanilla , Oaxaca, 31 December 2019).
Specimens examined: 8 specimens off Acapulco , Guerrero, (6°51 ′ N, 99°52 ′ W), collected by J. Violante-Gonzalez, July 2010 ( CNHE: 8287) GoogleMaps .
Site of infection: Wall of the pharyngeal cavity, gill arches, and top of the esophagus.
Prevalence: 2 of 2 hosts infected with a total of 19 worms.
ZooBank registration: urn:lsid:zoobank.org:act:DD82BB40-583D-47BE-8955-139832C5F321 .
Etymology: The species name refers to the pelagic habits of the host.
Representative DNA sequences in GenBank: partial 28S: OK018174 View Materials ; partial 18S: OK018175 View Materials .
Phylogenetic position of Peruanocotyle pelagica
The total lengths of the newly generated 18S and 28S sequences of Peruanocotyle pelagica are 1,590 and 1,288 nucleotides, respectively. Phylogenetic analysis based on the 28S locus recovered Monocotylidae as monophyletic; however, Dasybatotreminae , represented in the analysis by Troglocephalus rhinobatidis Young, 1967 , was recovered as polyphyletic because this species was found grouped with 3 species of Neoheterocotyle (Heterocotylinae) , whereas P. pelagica (member of Dasybatotreminae ) was found in a polytomy together with clades of Heterocotylinae, Loimoinae, and Monocotylinae ( Fig. 11 View Figure 11 ).
Remarks
The genus Peruanocotyle was erected by Chero et al. (2018) to accommodate specimens with a broadly ovoid anterior attachment organ (with numerous grooves and antero-medial notch); mouth surrounded by dome-shaped papillae; anchors shorter than the width of the marginal membrane; ceca with diverticula; MCO whip shaped, composed of 4 connate tubes, which end in 4 interlaced extensions, two of them armed; 2 pairs of testes and proximal ovary with clavate lobes. All these characteristics are present in the new species described here; however, P. pelagica can be differentiated from P. chisholmae on the combination of the following characters: lack of an accessory piece associated with anchors in P. pelagica ; presence of dendritic structures in marginal membrane of haptor, anchors, and hooklets embedded in individual membranous processes; radial and outer-rim septa with numerous haptoral papillae; a simple internal seminal vesicle, which is bipartite in P. chisholmae ; absence of an external seminal vesicle; a non-sclerotized vagina, which is sclerotized in P. chisholmae ; 5 spherical ovarian lobules, where P. chisholmae has 6; no accessory piece associated with the MCO, and 20 spines on each of the armed sections of the distal portion of the MCO, where those of P. chisholmae are only 14 or 15. Interlaced extensions of MCO end straight and connected in the new species, while extensions end curved and independent in the Peruvian species. Another potential difference between both species is the presence of anterior glands reported by Chero et al. (2018) for P. chisholmae , since these glands were not observed in P. pelagica ; however, some sort of glandular secretion associated with the muscular grooves was observed in SEM microphotographs ( Fig. 8 View Figures 4–8 ), suggesting that these glands may be present in the new species described here.
In a study by Violante-González et al. (2018) on the helminth community of R. steindachneri off Acapulco Bay, Guerrero, Mexico, 45 specimens of an unidentified monocotylid monogenean ( Dasybatotreminae gen. sp.) were reported attached to the gills of this ray species. Observations on eight of these specimens, 6 deposited in the CNHE under the number 8287 and 2 in the Colección de Morfologıá Animal (COMA), Instituto de Biologıá, Universidad Autónoma del Estado de Hidalgo (P64), confirmed that they possess the diagnostic characters of the species described herein, so we identify them as P. pelagica .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |