
Transversotrema damsella
|
publication ID |
https://doi.org/ 10.5281/zenodo.211252 |
|
DOI |
https://doi.org/10.5281/zenodo.6179408 |
|
persistent identifier |
https://treatment.plazi.org/id/74747F77-2262-FF95-FF44-FB0F15E7FF6A |
|
treatment provided by |
Plazi |
|
scientific name |
Transversotrema damsella |
| status |
|
Transversotrema damsella View in CoL n. sp
( Fig. 8 View FIGURE 8 )
Type-host: Abudefduf bengalensis (Bloch) (Pomacentridae) , Bengal Sergeant
Type-locality: Lizard Island, northern GBR, Queensland, Australia, (14°40’S 145°28’E)
Other hosts: Pomacentridae : Abudefduf septemfasciatus (Cuvier) , banded sergeant; Abudefduf sexfasciatus (Lacepède) , Scissortail Sergeant; Amblyglyphidodon curacao (Bloch) , Staghorn damselfish; Pomacentrus moluccensis Bleeker Lemon damsel. Labridae : Thalassoma hardwicke (Bennett) , Six-bar Wrasse. Mugilidae : Crenemugil crenilabis (Forsskål), Fringelip mullet.
Site: Beneath the scales
Material examined: Table 3 & 4. Neither Thalassoma hardwicke nor Pomacentrus moluccensis appear in the table because these specimens were given to us by Dr. G. Muñoz without associated prevalence data.
Molecular sequence data: ITS2 rDNA
GenBank accession numbers: Table 2
Deposited specimens: Holotype QMG 231854 (ex A. sexfasciatus LI coll. Cribb et al. 4 Apr 2006) and paratypes QMG 231855 (ex A. sexfasciatus LI coll. Cribb et al. 4 Apr 2006), QMG 231856 (ex A. septemfasciatus LI coll. Cribb et al. 4 Apr 2006), QMG 231857 (ex A. bengalensis LI coll. Cribb et al. Jun 2007), QMG 231858 (ex A. bengalensis LI coll. Cribb et al. Jun 2007).
Etymology: As the common name for pomacentrids is damsel fish the species name damsella was chosen for this species.
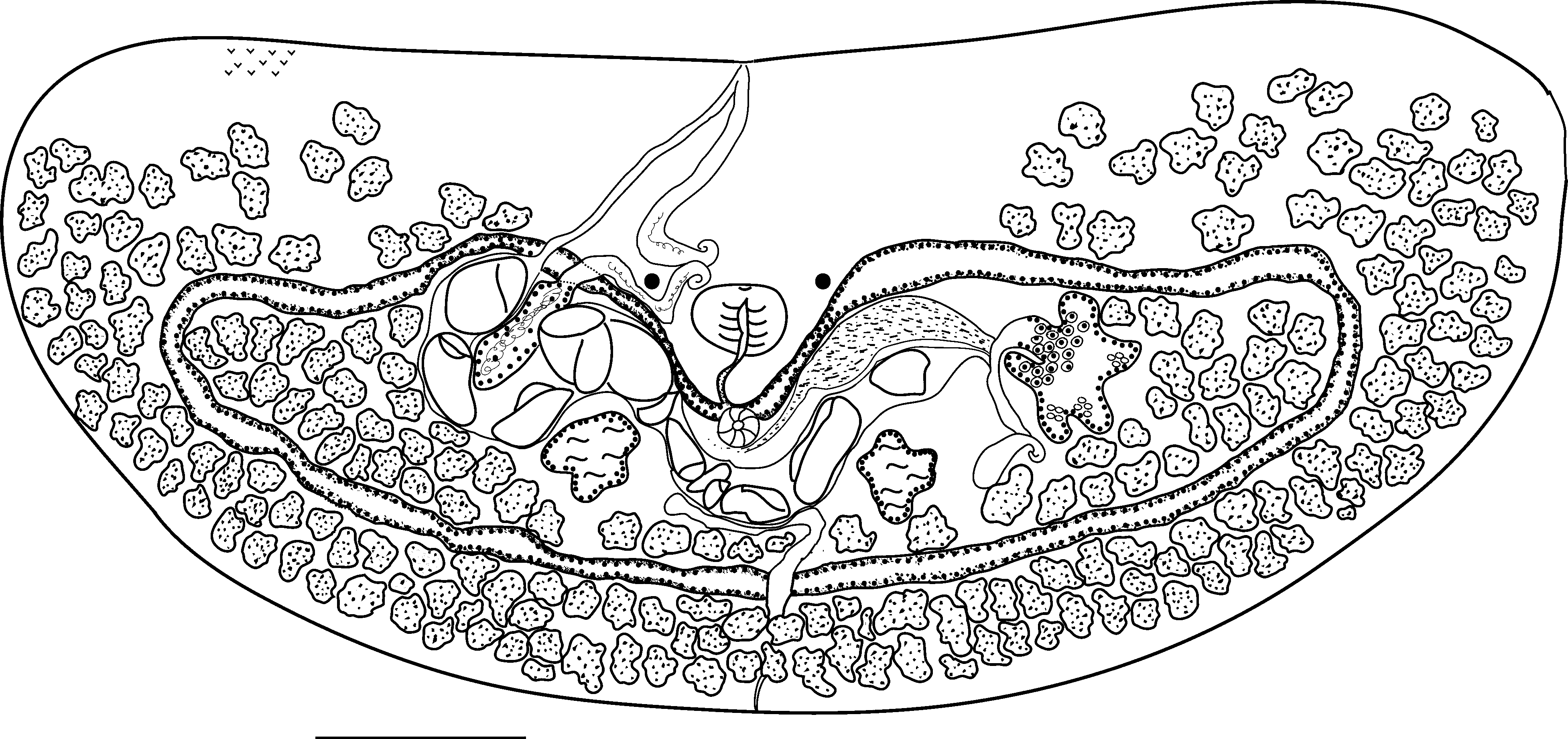
Description: Based on measurements of 6 specimens from pomacentrids from Lizard Island. Body D-shaped, strongly dorsoventrally flattened, 685–964 (808) long, 1503–2410 (2001) wide; average width/length range 2.5:1. Pharynx to anterior margin 203–375 (266); cyclocoel to posterior margin 107–141 (116). Tegumental spines prominent. Eyespots prominent 193–244 (214) apart, 9.7–12.8 (11.75%) of body width; no pigment evident other than in eyespots. Ventral sucker well posterior to eyespots, 3,022–6,878 (4,512) μm2. Mouth mid-ventral, inconspicuous. Pharynx slightly posterior to eyespots, 84–101 (93.7) long, 48–65 (53.25) wide. Oesophagus curved 79–236 (131). Caecal bifurcation dorsal to ventral sucker. Caeca-form cyclocoel reaching laterally to envelop testes, ovary and some vitelline follicles. Testes opposite, deeply lobed, left, 10,222–14,084 (12,444) μm2; right 10,863–13,653 (12,118) μm2. Seminal vesicle formed of lobed, saccular enclosed portion and winding, tubular extracaecal portion. Enclosed portion distinctly lobed or entire, antero-dextral to right of testis, constricts distally to form narrow duct that passes ventral to cyclocoel to join tubular portion. Tubular portion of seminal vesicle passes mediad along cyclocoel then turns anteriorly and passes between eyespots dextral to pharynx and passes to common genital pore where it unites with uterus without any specialisation. Common genital pore precisely in midline on anterior margin of worm. Ovary sinistral to left testis, prominent extended lobes, 9,051–18,342 (14,204) μm2, an average of 28.4% larger than testes. Oviduct passes medio-posteriorly, unites with Laurer’s canal and duct from oviduct passes vitelline reservoir, Laurer’s canal then passes posteriorly to open dorsally close to left testis; median portion dilated, contains sperm or vitelline remnants. Vitelline reservoir immediately anterior to left testis. Extracaecal vitelline follicles large, confluent, lateral and posterior to cyclocoel, loosely assembled anterior to cyclocoel from midway between excretory vesicle and lateral margins and extending to anterior margin. Enclosed follicles (av. 55) in two loosely assembled masses at each lateral extremity scattered posteriorly to testes along inner margins of cyclocoel. Uterus passes medially between anterior half of cyclocoel and testes then between right testis and saccular portion of seminal vesicle. Proximal portions of uterus act as seminal receptacle. Eggs tanned, 65–110 (85.3) long and 36–75 (49) wide, average number in utero 15. Excretory bladder opens posteriorly at small notch in middle of posterior margin, extends anteriorly in initially narrow tube which then expands into large sac which passes ventral to cyclocoel anterior to which it becomes laterally directed.
Remarks: Transversotrema damsella n. sp. is the most distinctive species from this complex. The body is Dshaped rather than crescent - shaped as in T. licinum and body proportions are different. T. damsella n. sp. has an ovary which is larger than the testes and this distinguishes it from all other species in the complex. The oesophagus is also much longer than those found in T. licinum , T. atkinsoni n. sp., T. borboleta n. sp. and T. carmenae n. sp. The vitelline follicles are large, scattered randomly to the lateral margins and in the anterior margin to approximately half way from the lateral margins to the genital pore. The enclosed follicles are also large and numerous and scattered posterior to the testes along the posterior edge of the cyclocoel. The small testes and large ovary, combined with the distribution of the follicles, both external to and enclosed by the cyclocoel, separates this species from all others. Although we have two clear specimens of this species from the labrid Thalassoma hardwicke donated by G. Munoz, we found no specimens of this species ourselves from examination of 73 labrids at Lizard Island. These fishes did not include T. hardwicke but eight individuals of two other species of Thalassoma were not infected.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |