
Chaetonotus (Chaetonotus) antrumus Kolicka, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.354 |
|
publication LSID |
lsid:zoobank.org:pub:51C2BE54-B99B-4464-8FC1-28A5CC6B9586 |
|
DOI |
https://doi.org/10.5281/zenodo.3851697 |
|
persistent identifier |
https://treatment.plazi.org/id/BF874601-C073-465F-B50B-48D73DB5A4C5 |
|
taxon LSID |
lsid:zoobank.org:act:BF874601-C073-465F-B50B-48D73DB5A4C5 |
|
treatment provided by |
Carolina |
|
scientific name |
Chaetonotus (Chaetonotus) antrumus Kolicka |
| status |
sp. nov. |
Chaetonotus (Chaetonotus) antrumus Kolicka View in CoL sp. nov.
urn:lsid:zoobank.org:act:BF874601-C073-465F-B50B-48D73DB5A4C5
Figs 4–11 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig ; Table 4 View Table 4 ; Appendix
Diagnosis
Slender body, length from 91.2 to 129.7 μm. Head five-lobed, cephalion and pleuria weakly marked in the head outline. Hypostomium small and rhomboidal. Ocellar granules absent. Scales small, threelobed and with strong keels. One pair of one-lobed, keeled scales on the dorsal side of the posterior part of the trunk, and on the dorsal and dorsolateral sides of the furcal appendages. Two pairs of three-lobed, spined scales present on the ventral side of the furcal appendages. Scales distributed in 29–35 total longitudinal rows (11–13D+6–8DL+6L+4–6LV+ 2V) with 23–27 scales in the central row, and differing morphologically in the areas of the head, neck and trunk, respectively. Spines thick, simple, with blunt ends. Spine lengths strongly vary: spines of the head are longer than those of the neck; spines of the neck are short, but become progressively longer to the widest body region, after which they gradually shorten towards the furcal base. The spines gradually increase in length from lateral to ventral body side towards the ciliary bands. Ventral scale spines longer than the others and hair-like. Last pair of parafurcal spines longer and stronger. Ventral interciliary field naked, except for the posterior trunk region. A pair of ventral terminal scales long, oval with shallow posterior notches, keeled and spineless. Pharynx narrow with two weakly marked dilatations. Straight intestine with a distinct, short anterior section appearing as a narrow band.
Etymology
From Latin ‘ antrum ’, cave, referring to the habitat where the species was found.
Material examined
Holotype
MONTENEGRO:adult, Crnojevica River flowing in Obodska Cave, 42°21.118′ N, 19°0.304′ E, sample 3, sampling site 3, 29 Jul. 2015, Piotr Gadawski leg., determined by Małgorzata Kolicka (photomicrograph in the Natural History Collections, Adam Mickiewicz University, Poznan: NHC-GCCA- 12-1-20 /h).
GoogleMapsParatypes
MONTENEGRO: 31 adults, 2 subadults, 2 juveniles, Crnojevica River flowing in Obodska Cave, 42°21.118′ N, 19°0.304′ E, samples 1 and 3, sampling sites 1 and 3, 29 Jul. 2015, Piotr Gadawski leg., determined by Małgorzata Kolicka (Natural History Collections, Adam Mickiewicz University, Poznan under accesing number: NHC-GCCA-12-21-50).
Description
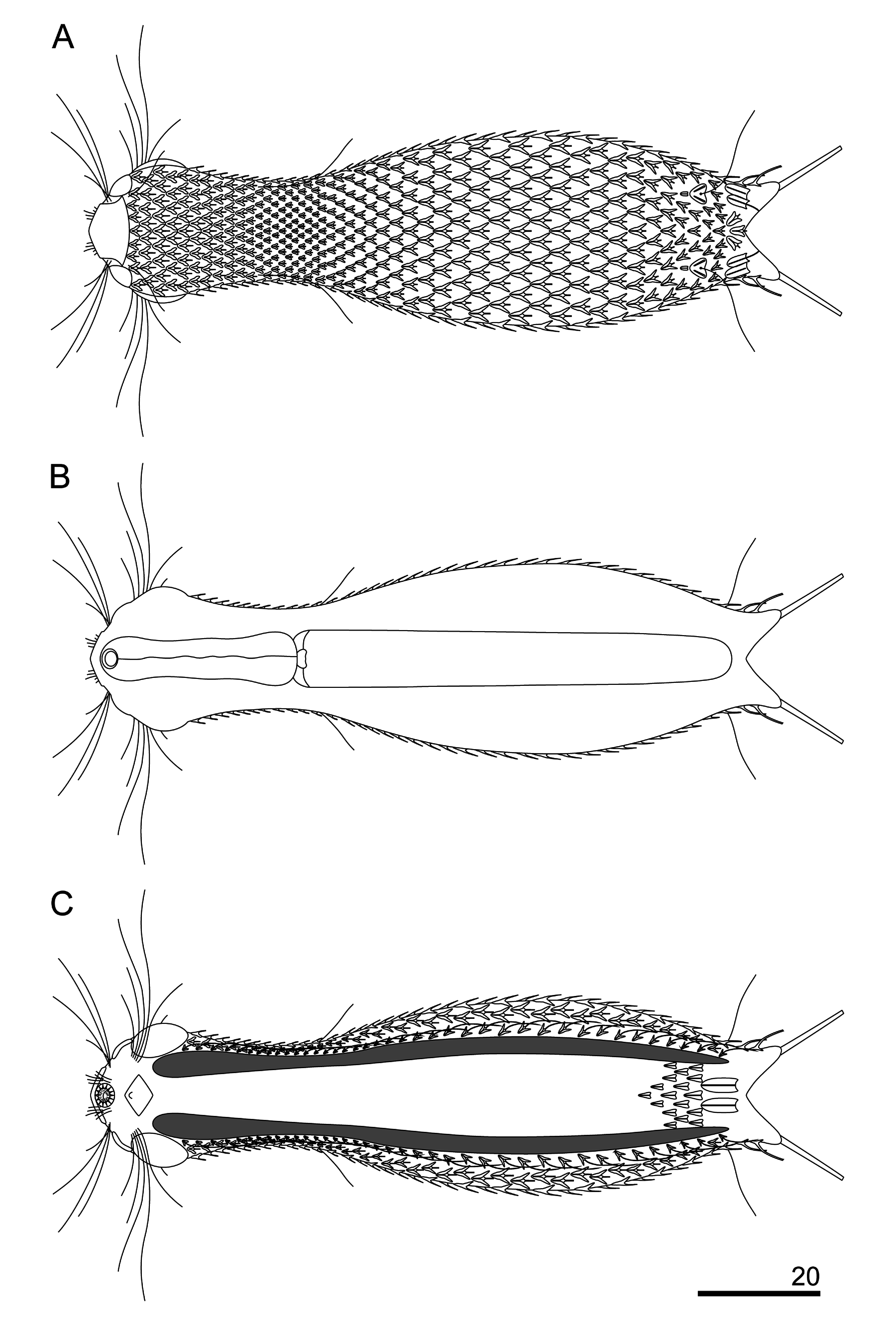
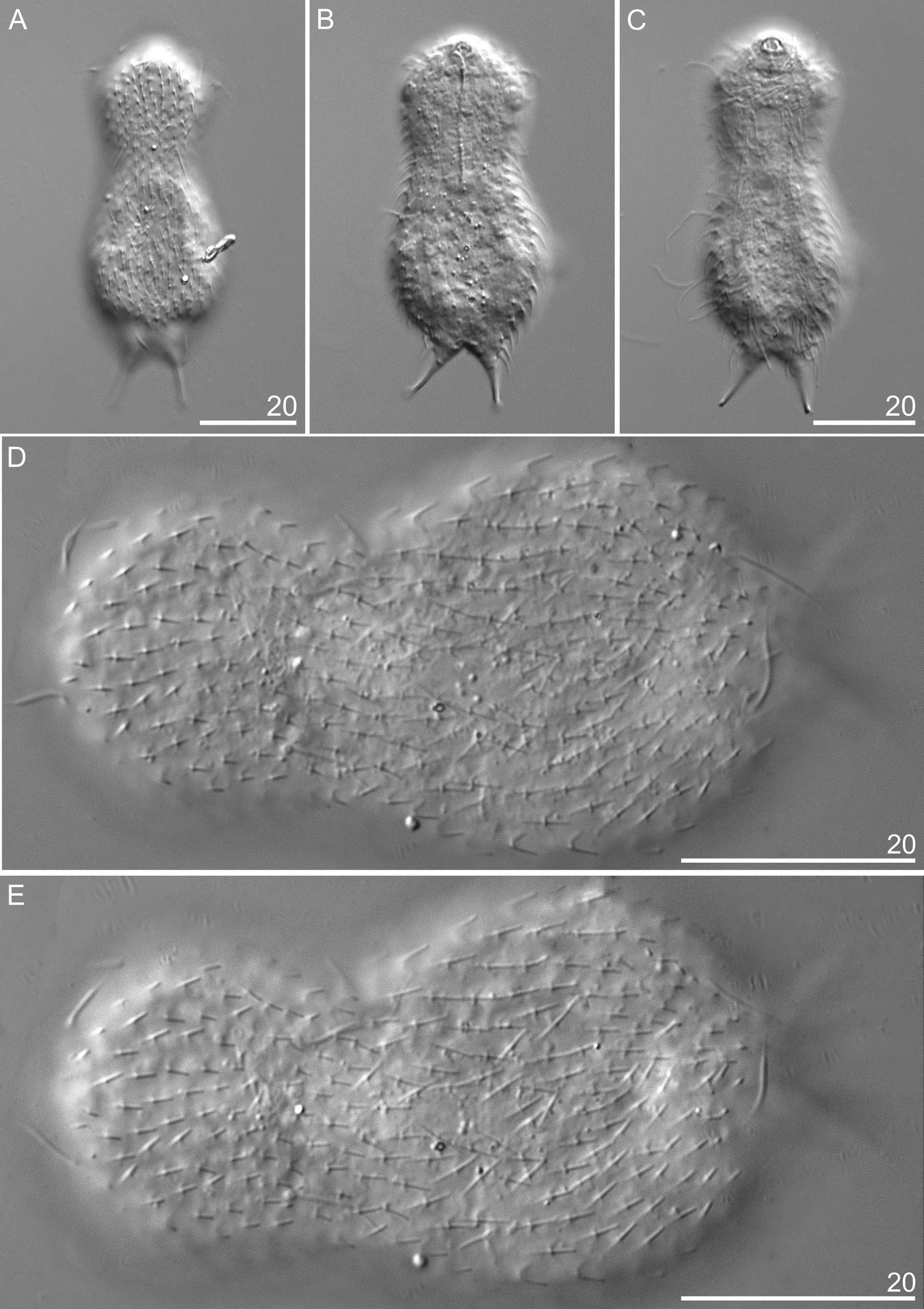
This new species has a slender body. Its head is wider than the neck and separated from the trunk by a distinct neck constriction. The neck extends into the trunk, which gradually widens towards its widest region beyond its midpoint (ca U61), after which it gradually tapers towards a distinct furcal base at U84. The branches of the furca are set wide apart. The furcal indentation is V-shaped, and the adhesive tubes diverge posteriorly. They measure 10.1–11.3 μm, are straight and thin and do not taper towards their blunt ends ( Figs 4–6 View Fig View Fig View Fig ).
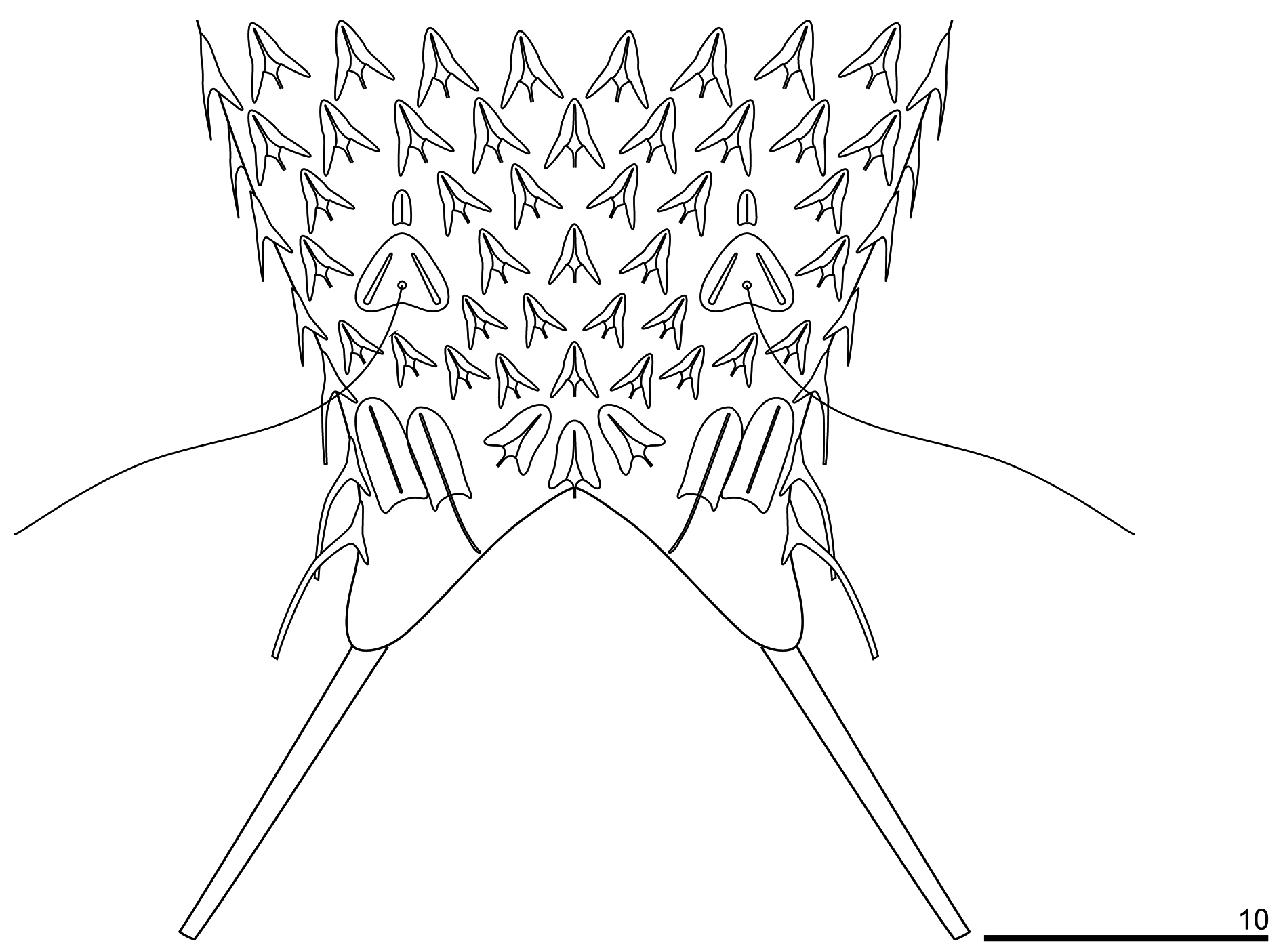
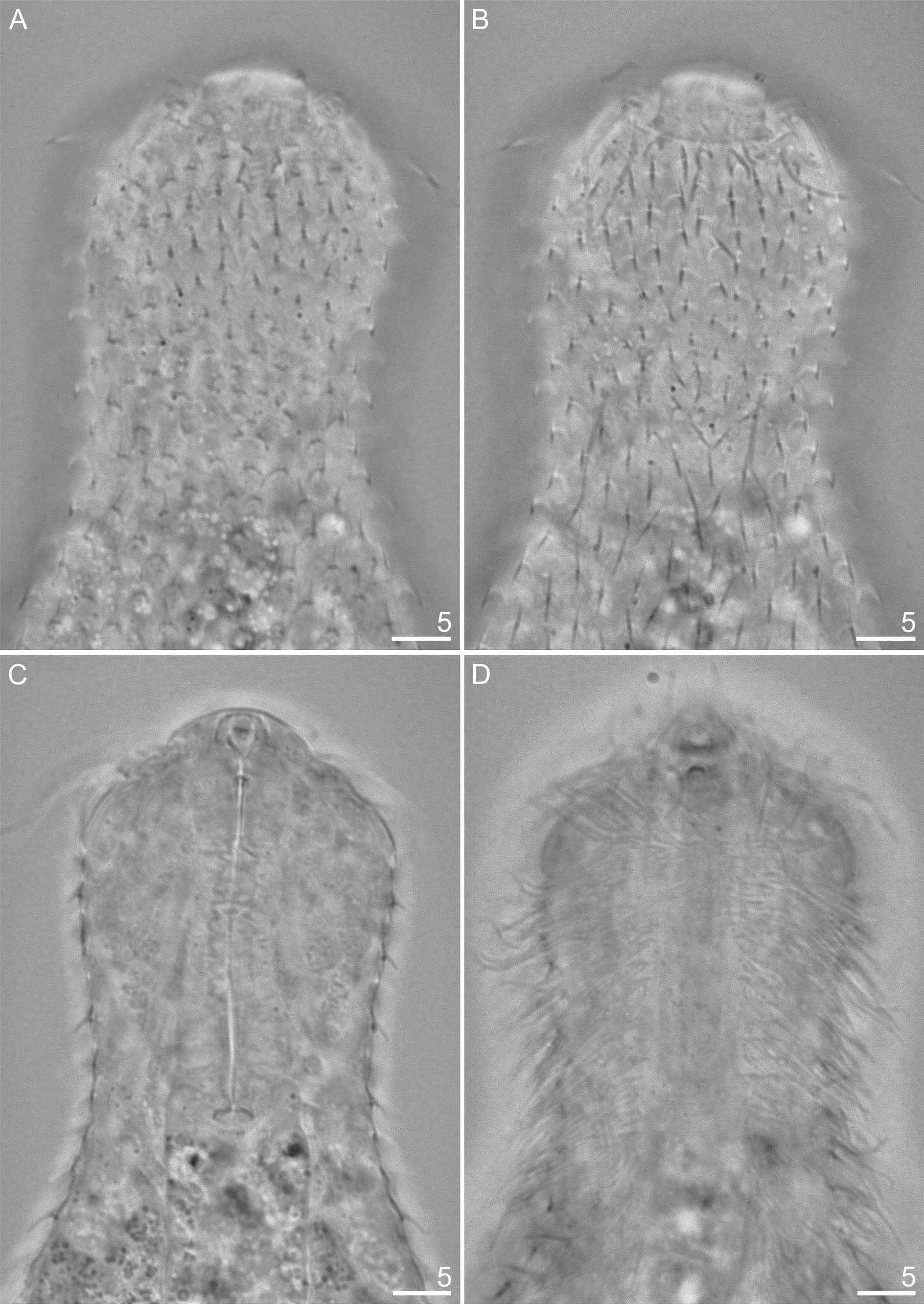
The head is five-lobed and semi-circular. All plates are visible in the dorsal head outline. The cephalion (U1–U5) adheres to the head along its entire length, is narrow and widens at the dorsal edge. The epipleuria (U4–U6) are small and slightly convex. The hypopleuria (U7–U13) are more than twice as large as the epipleuria. The hypopleuria are not visible from the dorsal side; only their outline is marked in body shape ( Figs 4A View Fig , 7 View Fig ). The hypostomium (U5–U8) is short and rhomboidal with slightly rounded edges and a strong anterior edge ( Fig. 4C View Fig ). Two pairs of cephalic ciliary tufts are present. The anterior tufts have four cilia each that emerge from between the cephalion and epipleuria and are arranged in lines around the lateral edges of the cephalion. The beginning of these lines (the first two cilia) is clearly visible on the dorsal side. The anteriormost cilium in both anterior tufts is fairly short (the shortest). The second cilium is longer than the first. The third cilium is very long and is the longest of all cilia in the tuft. The posteriormost cilium is shorter than the third and similar in length to the second cilium. The posterior tufts have five cilia each and emerge ventrally at the anterior edge of the hypopleuria. The length of the cephalic cilia in the posterior tufts increases from the anteriormost to the fourth cilium. The posteriormost cilium is similar in length to the first (see Appendix). Ocellar granules are not present. The mouth ring is narrow, located sub-terminally at U2–U3 and has weakly marked, granular reinforcements. Short inner hairs are present inside the mouth ring and suboral hairs are located around it.
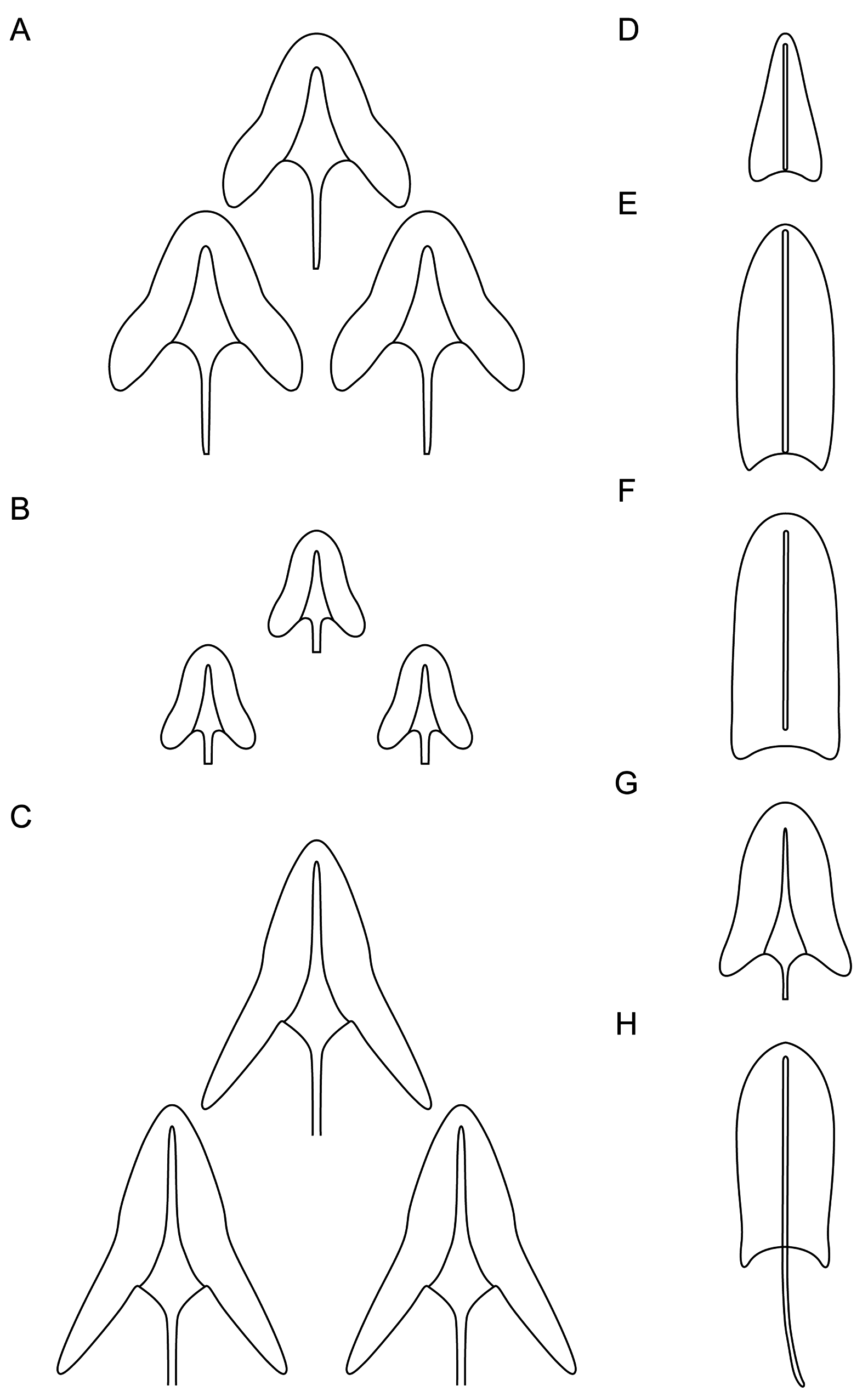
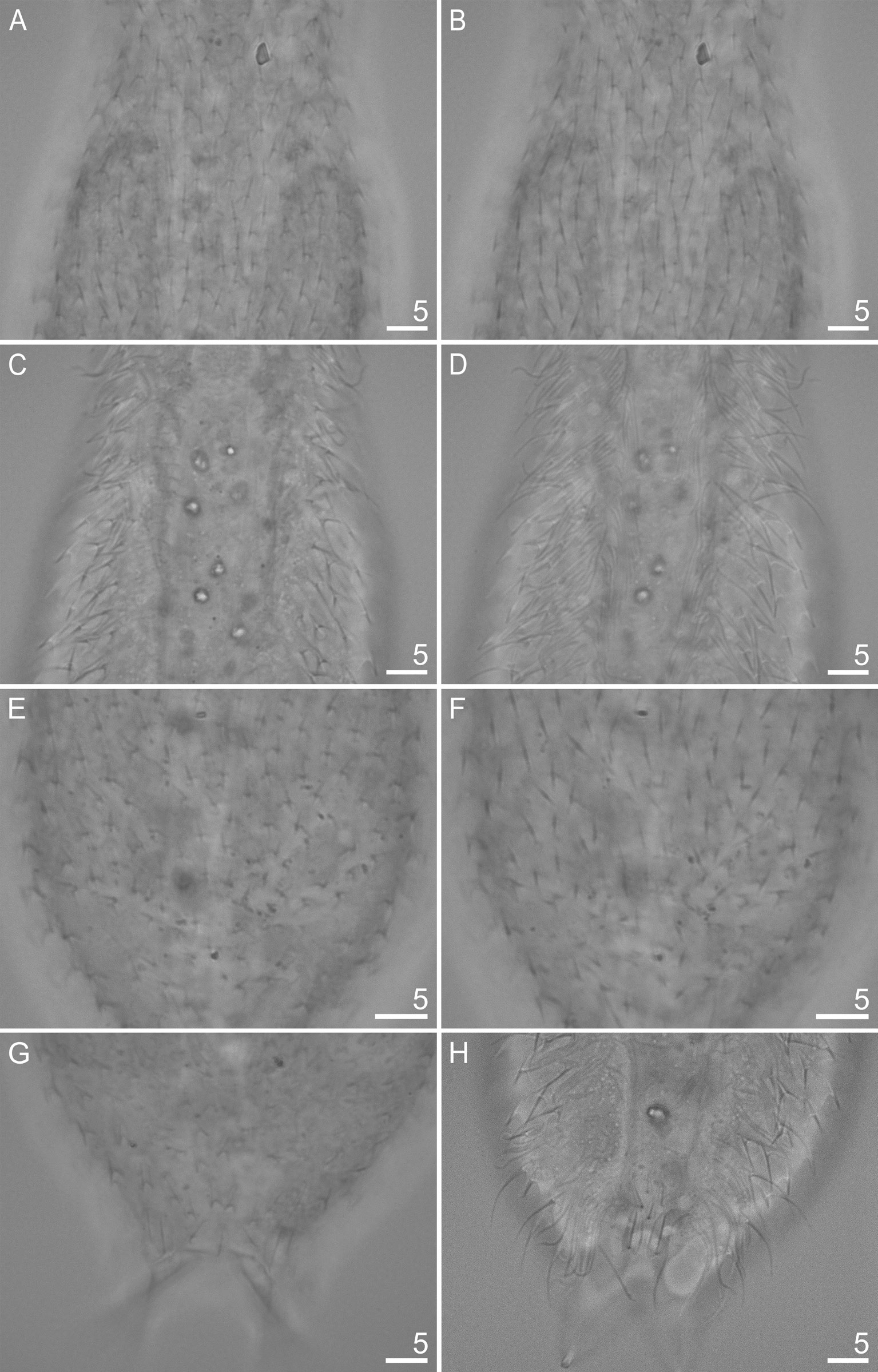
The body is covered by small, three-lobed scales and single one-lobed scales that adhere over their entire surface to the cuticle ( Figs 5–9 View Fig View Fig View Fig View Fig View Fig ). Scales are distributed in 29–35 total longitudinal and alternating rows (11–13D+6–8DL+6L+4–6LV+2V) with 23–27 scales in the dorsal central row. Each scale has a strong keel and triangular with a deeply-notched posterior edge. On the head, neck and anterior and middle parts of the trunk, the longitudinal rows of scales run parallel to each other, while in the posterior part of the trunk and on the furcal base the scales gradually converge towards the central longitudinal row ( Figs 4 View Fig , 7–9 View Fig View Fig View Fig ). The scales on the head, neck and trunk are arranged close to one another, but their edges do not adhere or overlap. The longitudinal rows of scales begin on the head beyond the posterior edges of the cephalion, epipleuria and hypopleuria. The scales show a strong morphological diversity in posterolateral lobe distinctness, edge roundness and size throughout the different surfaces of the head, neck and trunk regions. On the head, there are deeply-notched, rounded, wide triangles with rounded posterolateral lobes that are weakly differentiated from the central lobe. On the neck, scales are narrower, and their postero-lateral lobes show a weaker separation from the central lobe (see Appendix; Figs 4A View Fig , 5 View Fig ). On the trunk, scales are shaped like narrow, deeply-notched triangles with their postero-lateral lobes clearly differentiated from the central lobe and with edges that are less rounded ( Figs 4–5 View Fig View Fig , 8A, C, E View Fig , 9A View Fig ). The size of the scales decreases rapidly at the beginning of the neck, after which it gradually increases towards the beginning of the trunk (head: 1.6–3.8 μm length × 1.8–4.3 μm width vs neck: 1.3–3.3 μm length × 1.4–3.9 μm width vs trunk: 2.5–5.6 μm length × 1.6–4.3 μm width). The dorsal head and neck scales differ most from one another ( Figs 4A View Fig , 5 View Fig , 7A View Fig ), whereas in the other areas of the head and neck, the differences in shape and size between the scales are gradual. The size of the scales of the trunk increases from the anterior end towards the widest trunk region, after which it decreases towards the furcal base (see Appendix). On the posterior part of the trunk, scales are clearly smaller than those at the widest point of the body. A fine (1.9–2.7 μm length × 1.3–1.8 μm width) one-lobed, keeled and spineless scale is located at U78 on the dorsal part of the trunk, anterior to each scale bearing a sensory bristle ( Figs 4A View Fig , 8C View Fig ) and is shaped like a strongly-rounded triangle. Medially, on the dorsal side of the furcal appendages (U86–U88), three three-lobed scales are present that are slightly narrower than the other scales of the trunk (see Appendix; Figs 4A View Fig , 8G View Fig ). Lateral to these scales, on the furcal appendages (U85–U88), there are two pairs of elongated one-lobed scales shaped like narrow ovals with a weakly-notched posterior edge. The anterior one of each pair of scales is located dorsally, has a strong keel and a long, straight spine, whereas the posterior one is located dorsolaterally and has a long keel, but no spine. The lateral edges of these scales are partially overlapping. On the lateral side of the furcal appendages (U86–U90), two pairs of three-lobed scales, of the same type as the scales of the trunk, with spines are present ( Figs 6 View Fig , 9A View Fig ). The dorsal, dorsolateral, lateral, ventrolateral and ventral scales do not strongly vary in size, except that the scales in the area of the neck and in the longitudinal rows next to the ciliary bands are considerably smaller than the others. The ventral scales of the longitudinal rows located closest to the ventral ciliary bands are about half the size of the scales in the other rows (head: 1.6–2.9 μm length × 1.8–2.3 μm width; neck: 1.3–1.8 μm length × 1.4–2.0 μm width; trunk: 2.5–3.0 μm length × 1.6–2.0 μm width) and have their central lobe rotated about 20° towards the bands (see Appendix; Figs 4C View Fig , 8C View Fig , 10C View Fig ).
The spines arising from the posterior scales region are thick and straight, taper very slightly towards their blunt ends and have no lateral denticles ( Figs 4 View Fig , 7B, D View Fig , 8B, D, F View Fig , 9B View Fig , 10E View Fig ). The spines that adhere directly to the cephalion and pleuria are the shortest of the head spines ( Fig. 7B View Fig ). The spines on the head increase in length (1.1–3.1 μm), whereas those on the neck decrease rapidly in length. The spines on the neck are much shorter than those in the head area; the spines are merely vestigial halfway down the neck, after which they gradually lengthen towards the trunk (0.5–2.7 μm) ( Figs 4 View Fig , 7B View Fig ). The spines on the trunk gradually and slightly lengthen from the beginning of the trunk (ca U30) up to the widest body region (ca U61), after which they gradually shorten towards the furcal base at U84 (1.3–4.1 μm) ( Figs 4 View Fig , 8B, D, F View Fig ). The pair of posteriormost lateral trunk spines is slightly longer and thicker than the surrounding spines (3.4–5.9 μm). Parafurcal spines emerging from two lateral scales per side on the furcal appendages are slightly thicker and longer than the other spines of the body, the posteriormost pair is longer, thicker and stronger than those of the preceding pair (see Appendix; Fig. 4 View Fig ). These spines taper slightly towards their blunt distal ends. The dorsal and dorsolateral spines do not vary substantially in length ( Table 4 View Table 4 ). The spines lengthen gradually and slightly from the lateral side towards the ciliary bands ( Fig. 9B View Fig ). The spines arising from the ventral longitudinal row of scales located closest to the ciliary bands are much longer than those of the body, curved, and hair-like along their entire length (head: 5.1–8.6 μm; neck: 5.0–9.0 μm; trunk: 7.1–14.0 μm).
This species has three pairs of dorsal sensory bristles ( Fig. 4 View Fig ). The first pair is located on the head, directly posterior to the cephalion, near the lateral edges of the epipleuria (U5), and each bristle emerges from a small, round papilla. The second pair is located on the neck (U25) and each bristle emerges from a small, rounded papilla. The third, posterior pair, which emerges from double-keeled scales located in
the posterior part of the trunk (U79–U80), is shaped like deeply-notched triangles with unconnected keels ( Figs 4A View Fig , 5 View Fig ).
On the ventral side, the longitudinal ciliary bands begin at U8 and run back to U84 ( Fig. 4C View Fig ). They are wider in the area of the head than in the other parts of the body. Most of the ventral interciliary field is naked: fine, keeled, spined scales are present only in the posterior part of the trunk (from ca U78 to U82) ( Figs 4C View Fig , 8H View Fig ). Their differentiation increases towards the posterior body region: the anterior scales are weakly delineated and partially submerged in the cuticle. These scales are shaped like narrow triangles with a very weakly notched posterior edge. The scales of the ventral interciliary field increase in size towards the posterior end of the body (1.4–3.7 μm length × 1.1–1.9 μm width). The terminal scales are located at U82–U85 and are shaped like long, narrow ovals with a very weakly notched posterior edge and have a long keel running along their entire length, but are spineless ( Figs 4C View Fig , 5E View Fig ).
The pharynx (U2–U28) is relatively narrow and has weak anterior and posterior dilatations, with the posterior dilatation wider than the anterior one ( Figs 4B View Fig , 7C View Fig ; Appendix). The pharynx connects through the small and narrow pharyngeal intestinal junction (U30) to a straight intestine (running from U29 to U86). The intestine has a distinct, short (U29–U31) anterior section marked as a narrow band ( Fig. 4B View Fig ).
Remarks
Chaetonotus (Chaetonotus) antrumus sp. nov. is an interstitial species which was recorded in the lotic system in a cave. The gastrotrich fauna in interstitial freshwater habitats is relatively rich, but composed of fewer species than in epibenthic or periphytic habitats (e.g., Balsamo et al. 2015). Interstitial communities are composed of not only taxa specific to them, but also of eurytopic species. Out of ca 40 species found in sandy biotopes, fewer than 10 seem to constitute exclusively interstitial taxa (Balsamo et al. 2015). All of the species share certain morphological traits, e.g., a small body size, a poorly ornamented cuticular covering, a well developed locomotory ciliature and adhesive organs ( Balsamo & Fregni 1995; Balsamo et al. 2015). Entire sets of these characteristics also occur in C. (C.) antrumus sp. nov.
Usually, one or two pairs of sensory bristles have been described in Chaetonotidae , but in C. (C.) antrumus sp. nov., three pairs of dorsal sensory bristles are present. The presence of dorsal sensory bristles in the area of the head has previously been noted in only two species of Chaetonotus : C. (C.) brevispinosus Zelinka, 1889 and C. (C.) sanctipauli Kisielewski, 1991 . Perhaps three pairs of dorsal sensory bristles occur more frequently in Chaetonotidae ; the location of the first pair of dorsal sensory bristles on the head near the cephalion and the cephalic cilia, however, may have led to these sensory bristles being obscured by the cilia or erroneously interpreted as one of the cilia. Thus, the presence of dorsal sensory bristles on the head cannot be considered as a good diagnostic characters at the species level.
0
.
04
The presence of a developing egg was observed in 21 out of 32 adult specimens of C. (C.) antrumus sp. nov. However, sperm and an X-organ were not observed.
Chaetonotus (Chaetonotus) antrumus sp. nov. showed very quick locomotion under microscopic observation. The species often remained in motion even after its integument had burst from compression under a cover glass, thus producing a visible deformation.
Sequence diversity and phylogenetic relationships
The COI alignment for the distance calculations comprised 628 bp of unambiguous sequence data for three specimens of Chaetonotus (Chaetonotus) antrumus sp. nov. We found two haplotypes that differed in eight nucleotide positions (K2P = 0.013; SD = 0.005). All substitutions were located at synonymous sites, i.e., both haplotypes coded for the same amino acid sequence. The nuclear data, including 4809- bp of the DNA region coding for 18S rRNA, ITS1, 5.8S, ITS2, and 28S rRNA, showed no intraspecific variation. All sequences are deposited in GenBank under accession numbers KU705230 View Materials , KU705231 View Materials (COI); KX538804 View Materials (18S); KU705232 View Materials (28S); KU705233 View Materials (ITS).
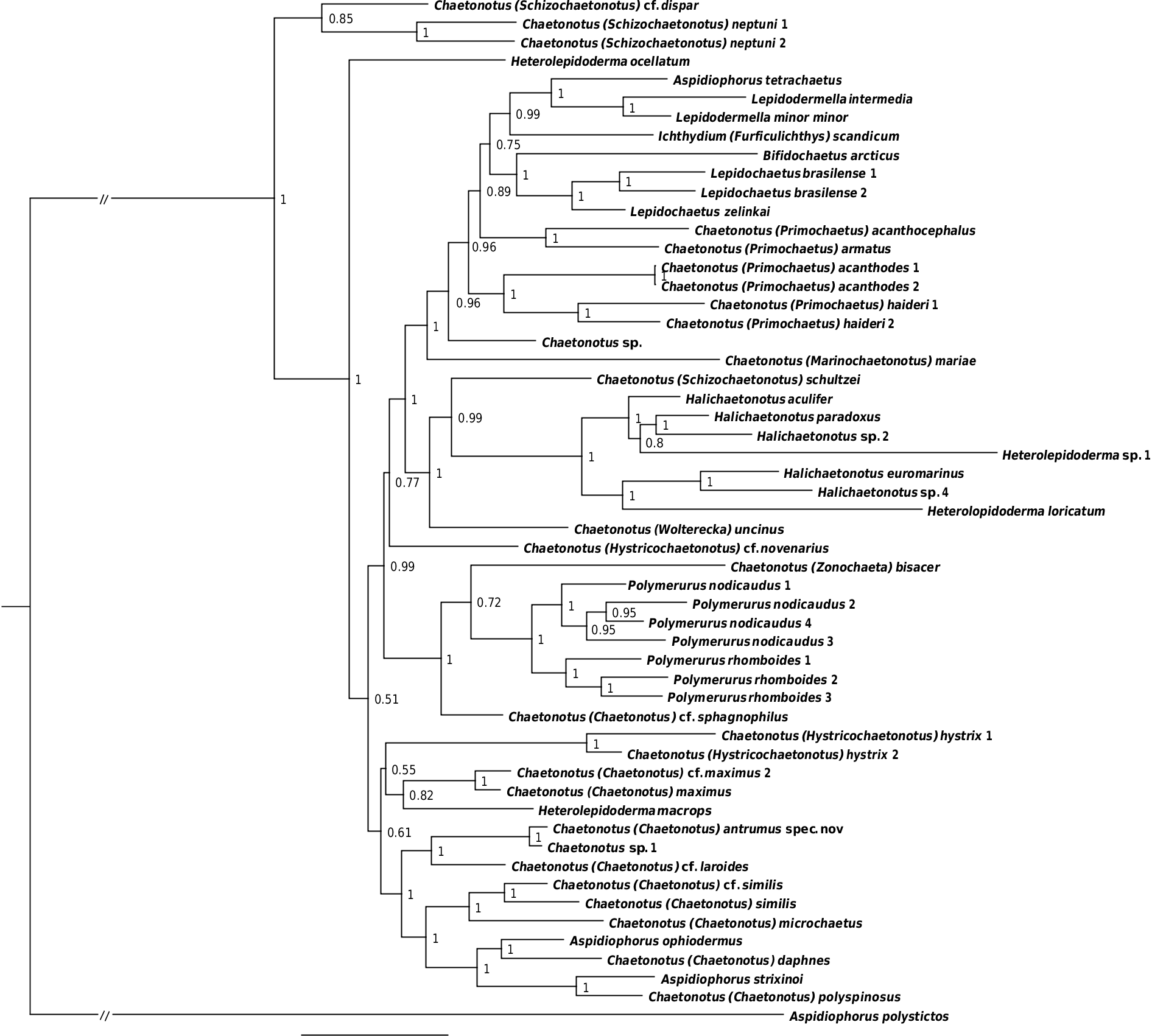
A phylogenetic analysis ( Fig. 11 View Fig ) recovered, with maximum support, C. (C.) antrumus sp. nov. as closely related to the undetermined Chaetonotus sp. 1 TK-2012 ( Kånneby et al. 2013); both grouped with C. (C.) cf. laroides. This clade was in a sister relation to a clade consisting of some representatives of Chaetonotus (Chaetonotus) Ehrenberg, 1830 and Aspidiophorus (Voigt, 1903) and including Chatonotus (Chaetonotus) daphnes Balsamo & Todaro, 1995 , which is morphologically most similar to the new species.
Differential diagnosis
Even if the new species was recorded from a sandy habitat, it is not similar to any other species considered as exclusively interstitial taxa. Chaetonotus Ehrenberg, 1830 is a polyphyletic genus, containing varied species. Of all the 165 currently known nominal freshwater representatives of this genus, C.(C.) antrumus sp. nov. is morphologically closest to C. (C.) naiadis Balsamo & Todaro, 1995 and C. (C.) daphnes Balsamo & Todaro, 1995 , both of which were originally reported from Italian mountain pools. These two species were selected from all the Chaetonotus representatives for the comparison of the new species due to a similarity in terms of (1) the alignment of scales whose edges are juxtaposed; the presence of a different type of scales on the dorsal and dorsolateral sides of the furcal base; (2) the type of scales on the ventral interciliary field; (3) the number of terminal scales of the ventral interciliary field; (4) two longitudinal bands of ventral locomotor cilia wider on the head region. Despite the fact that from all of the hitherto known species, C. (C.) naiadis and C. (C.) daphnes have the highest number of common features with the newly described species, they are significantly different from C. (C.) antrumus sp. nov., most strikingly in the scale type and shape. All the differences between the new species and the most morphologically similar taxa have been summarized in Table 4 View Table 4 .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |