
Hadrosaurs —
|
publication ID |
https://doi.org/ 10.5281/zenodo.3614972 |
|
DOI |
https://doi.org/10.5281/zenodo.4420215 |
|
persistent identifier |
https://treatment.plazi.org/id/794487B6-D624-860E-FF47-F8D2FC82F9CC |
|
treatment provided by |
Jeremy |
|
scientific name |
Hadrosaurs — |
| status |
|
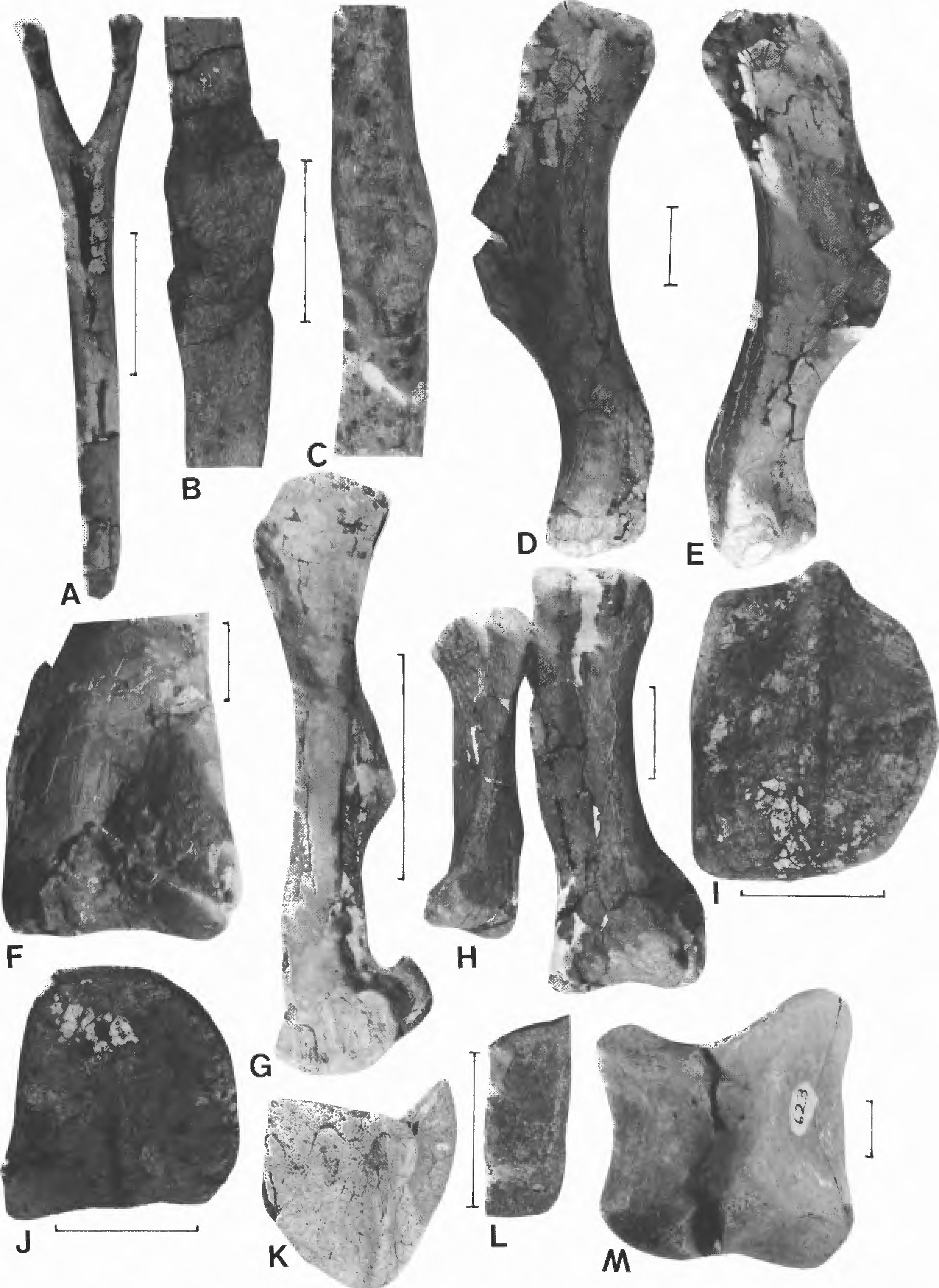
Several partial skeletons of hadrosaurs have recently been recovered from the Fruitland and Kirtland Formations. One such specimen from the Naashoibito Member was recently described by Hunt and Lucas (1991). A second partial skeleton is of an indeterminate hadrosaur from the upper Fruitland Formation of Ah-shi-sle-pah Wash ( Fig. 8 View FIGURE A-C, F-H). This specimen is interesting in that two '?successive ribs show healed fractures at about their midpoint ( Fig. 8 View FIGURE B-C).
A third hadrosaur skeleton from the Naashoibito Member of the Kirtland Formation includes much of a skull ( Fig. 9 View FIGURE 9 ) and associated postcranial elements ( Fig. 8 View FIGURE D-E). This specimen represents a hadrosaurine, as it possesses an anterodorsal process on the maxilla ( Homer, 1990) and has a flat dorsal skull profile (skull roof not illustrated). The most diagnostic element is the jugal ( Fig. 9 View FIGURE 9 A-D). The rounded lower margin of the lateral temporal fenestra, the narrow elongate postorbital process and the nearly right angle formed by the ventral margin below the lateral temporal fenestrae are of taxonomic value. In all these char acters, NMMNH P-1041 differs from Judithian and Edmontonian taxa and from the Lancian hadrosaurines Anatotitan (Weishampel and Horner, 1990, fig. 26.5C), Edmontosaurus regalis ( Lull and Wright, 1942, fig. 52) and E. annectens ( Lull and Wright, 1942, fig. 53). However, the new specimen closely resembles the jugal of Edmontosaurus saskatchewanensis ( Lull and Wright, 1942, fig. 56; Weishampel and Horner, 1990, fig. 26.5a) and we tentatively assign NMMNH P- 1041 to this taxon.
Fruitland and Kirtland hadrosaurs have been most recently placed in three species, Kritosaurus navajovius , Parasaurolophus tubicen and P. cyrtocristatus ( Lucas et al., 1987) . Recently it has been suggested that Kritosaurus is either a junior subjective synonym of Hadrosaurus or Gryposaurus (e.g., Chapman and Brett-Surman. 1990) or that the genoholotype of Kritosaurus is indeterminate (Weishampel and Horner, 1990). The genoholotype of Kritosaurus ( AMNH 5799 View Materials ) is a poorly preserved skull that is truncated at the anterior margin of the orbits. The entire lower jaw is preserved. We believe that this specimen can be distinguished from all other hadrosaur taxa and forms a basis for a distinct genus. Thus, Kritosaurus can be distinguished from most other hadrosaurine hadrosaurs (e.g., Edmontosaurus, Anattitan, Maiasaura , Prosaurolophus, Saurolophus, Shantungosaurus ) by the high, short lateral profile of the skull. Kritosaurus is distinguished from Brachylophosaurus by the lack of a posterior extension of the nasals dorsal to the orbit, from Aralosaurus by the possession of much larger lateral temporal fenestrae and from Hadrosaurus by differences in the ilium (see below). We consider Kritosaurus to be a valid genus and a sub jective senior synonym of Gryposaurus , contra Weishampel and Homer (1990).
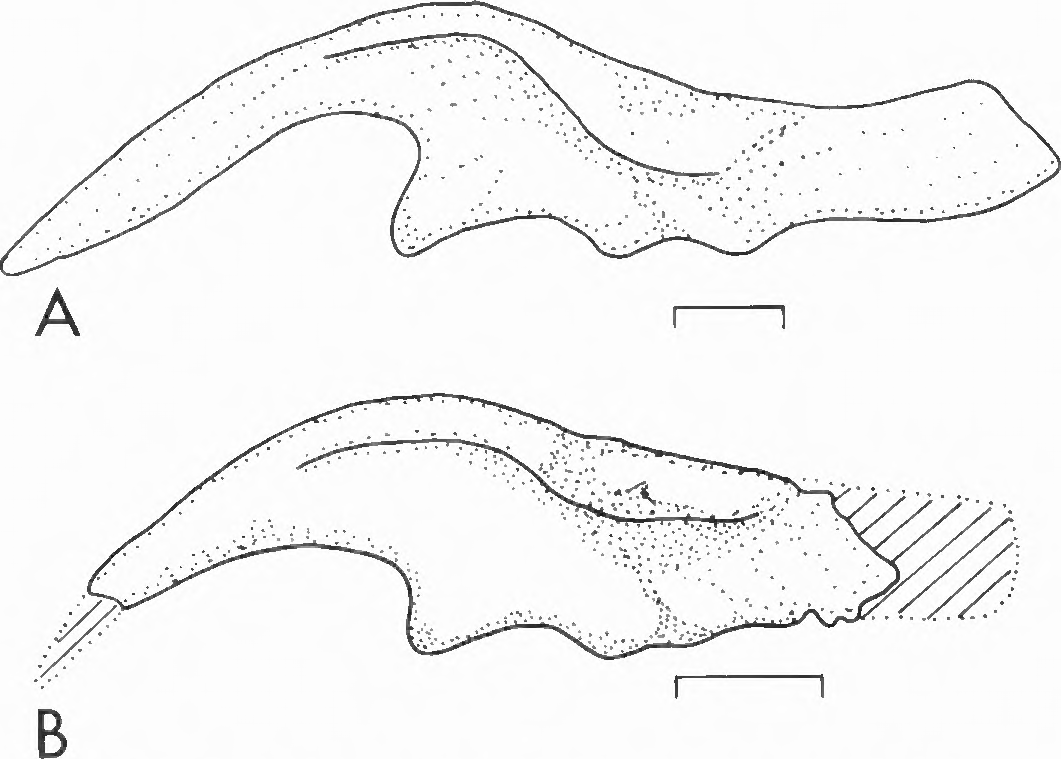
Hadrosaurus foulkii is based on a partial postcranial skeleton and fragments of a skull ( Lull and Wright, 1942, figs. 45-50). Only the postcranial specimens are complete enough for comparison with ma terial of Kritosaurus ( Lull and Wright, 1942; Pinna, 1979). The ilium is the most distinct of the elements represented in the skeleton of Hadrosaurus . Comparison of the ilia of Hadrosaurus foulkii (Leidy, 1856; Lull and Wright, 1942) and Kritosaurus navajovius ( Parks, 1920) indicates that there are significant differences between the two taxa ( Fig. 10 View FIGURE 10 ). The ilium of Hadrosaurus is distinguished principally by having a deeper and more abruptly tapering anterior process ( Fig. 10 View FIGURE 10 ). Davies (1983) noted the following other differences between the ilia of Hadrosaurus and Kritosaurus : (1) the antitrochanter is weak with little or no ventral deflection in Hadrosaurus , whereas in Kritosaurus the antitrochanter is robust with a strong ventral deflection; (2) the ridge extending anteriorly on the dorsal margin is weak with a short, straight extension onto the posterior process in Hadrosaurus , but in Kritosaurus the ridge is well developed and extends anteriorly on the dorsal margin; (3) in Hadrosaurus , posterior end of the antitrochanter is midway down onto the body of the ilium with a faint ridge extending dorsocaudally onto the posterior process; and (4) the ratio of acetabular length/height is less in Hadrosaurus (0.82) than Kritosaurus (0.92). These differences are consistent with the variation between genera rather than within genera (cf. Brett-Surman, 1975). We conclude that Kritosaurus is not a junior subjective synonym of Hadrosaurus .
We consider that there are no significant differences between Kri tosaurus navajovius and Kritosaurus notabilis and that they are synonyms. Kritosaurus breviceps (based on a partial dentary) and Kritosaurus marginatus (based on fragmentary postcrania) were assigned to the genus by Lull and Wright (1942) but we consider them nomina dubia . Kritosaurus incurvimanus is based on the posterior part of a skull and lower jaws and exhibits no differences from K. navajovius . Thus, we consider Kritosaurus to be a monospecific genus consisting only of K. navajovius .
Brown (in Sinclair and Granger, 1914) identified a maxilla and in complete dentary ( AMNH 5797 View Materials ) from the Naashoibito Member of the Kirtland Formation as belonging to Kritosaurus . However, these elements are not diagnostic at the generic level. Lucas et al. (1987) iden tified a right lower jaw of a hadrosaur from the Naashoibito Member as Kritosaurus ( Lucas et al., 1987) . This specimen is also generically indeterminate. Thus, there is no evidence for the presence of the had rosaurine Kritosaurus in the Naashoibito Member of the Kirtland Formation.
Two of the three species of Parasaurolophus , P. tubicen and P. cyrtocristatus , have been named for specimens from the Fruitland and Kirtland Formations. It is significant that the only three good skulls of Parasaurolophus were all named as different species (cf. Lucas, 1991). However, pending a revision of this genus, we consider both the San Juan Basin species to be valid. Hopson (1975) and Weishampel (1981) considered that the long crested P. walkeri and P. tubicen represent males and the short crested P. cyrtocristatus a female. Given that Parasaurolophus is rare in all faunas in which it occurs, is only known from three good skulls from three different stratigraphic units and that none of the three species co-occur, we consider speculations about sexual dimorphism in this genus to be unwarranted.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.