
Perissonemia occasa Drake, 1942
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.024 |
|
publication LSID |
lsid:zoobank.org:pub:C9003335-99FA-427E-988A-4A3CC2A73E4A |
|
DOI |
https://doi.org/10.5281/zenodo.10552979 |
|
persistent identifier |
https://treatment.plazi.org/id/7B372002-FF99-0514-044F-5EC1FBB213AF |
|
treatment provided by |
Felipe |
|
scientific name |
Perissonemia occasa Drake, 1942 |
| status |
|
Perissonemia occasa Drake, 1942 View in CoL
( Figs 1A, B View Figs 1 , 2A, B View Figs 2 , 3A View Figs 3 , 4A View Figs 4 , 5A View Figs 5 , 6A View Figs 6 , 7A View Figs 7 , 8A, D View Figs 8 , 9A, D View Figs 9 , 10A, B View Figs 10 )
Perissonemia occasa Drake, 1942:2 View in CoL . Holotype:♂, Japan:“3/14 Harima” [= Honshu, southwestern part of Hyogo-ken, iii.1914] (USNM) (see UNITED STATES NATIONAL MUSEUM OF NATURAL HISTORY 2021).
Perissonemia occasa: TAKEYA (1951) View in CoL : 18 (checklist: Japan); DRAKE & RUHOFF (1961): 135 (distribution); TAKEYA (1962): 56 (distribution); DRAKE & RUHOFF (1965): 324 (catalog); JING (1981): 311 (monograph); NAKANO (1984): 482 (host plant); MIYAMOTO & YASUNAGA (1989): 167 (checklist: Japan); TAKAHASHI (1990): 3 (checklist: Hyogo Prefecture); TSUKADA (1995): 49 (biology); P ÉRICART & GOLUB (1996): 53 (catalog: Palaearctic); YAMADA & TOMOKUNI (2012): 198 (monograph); YAMADA & ISHIKAWA (2016): 432 (checklist: Japan).
Baeochila occasa: LEE (1969) : 233 (male genitalia).
Material examined. Non-types (23 ♂♂ 22 ♀♀): JAPAN: HONSHU: Gifu-ken, Nakatsugawa-shi, Shizumo-yama, 7.x.2018, leg. Y. Yazaki (1 ♀, TUA); Mie-ken, Tsu-shi, Misugi-cho, Shimonogawa, 18.–21.iv.2011, leg.T. Shimada & Y. Hirano (3♀♀, TUA); Kyoto, Kitashirakawa, Kyoto Univ., 28.vii.1994, leg. M. Tsukada (4 ♂♂ 4 ♀♀, NSMT – referring to TSUKADA 1995); Prov. Harima [= southwestern part of Hyogo-ken], 2.vii.1906, leg. S. Iguchi (1 ♂ 1 ♀, ELKU – referring to TAKEYA 1962); Kobe [= Hyogo-ken, Kobe-shi], Oshibedani, Kizu, 8.viii.1982, leg. K. Nakano (1♀, NSMT – referring to NAKANO 1984); Hyogo-ken, Kobe-shi, Suma-ku, Ote, 2.viii.2020, leg. M.Yamashita (4 ♂♂, TUA), as above but 7.vii.2021, leg. J. Souma (14 ♂♂ 12 ♀♀, TUA).
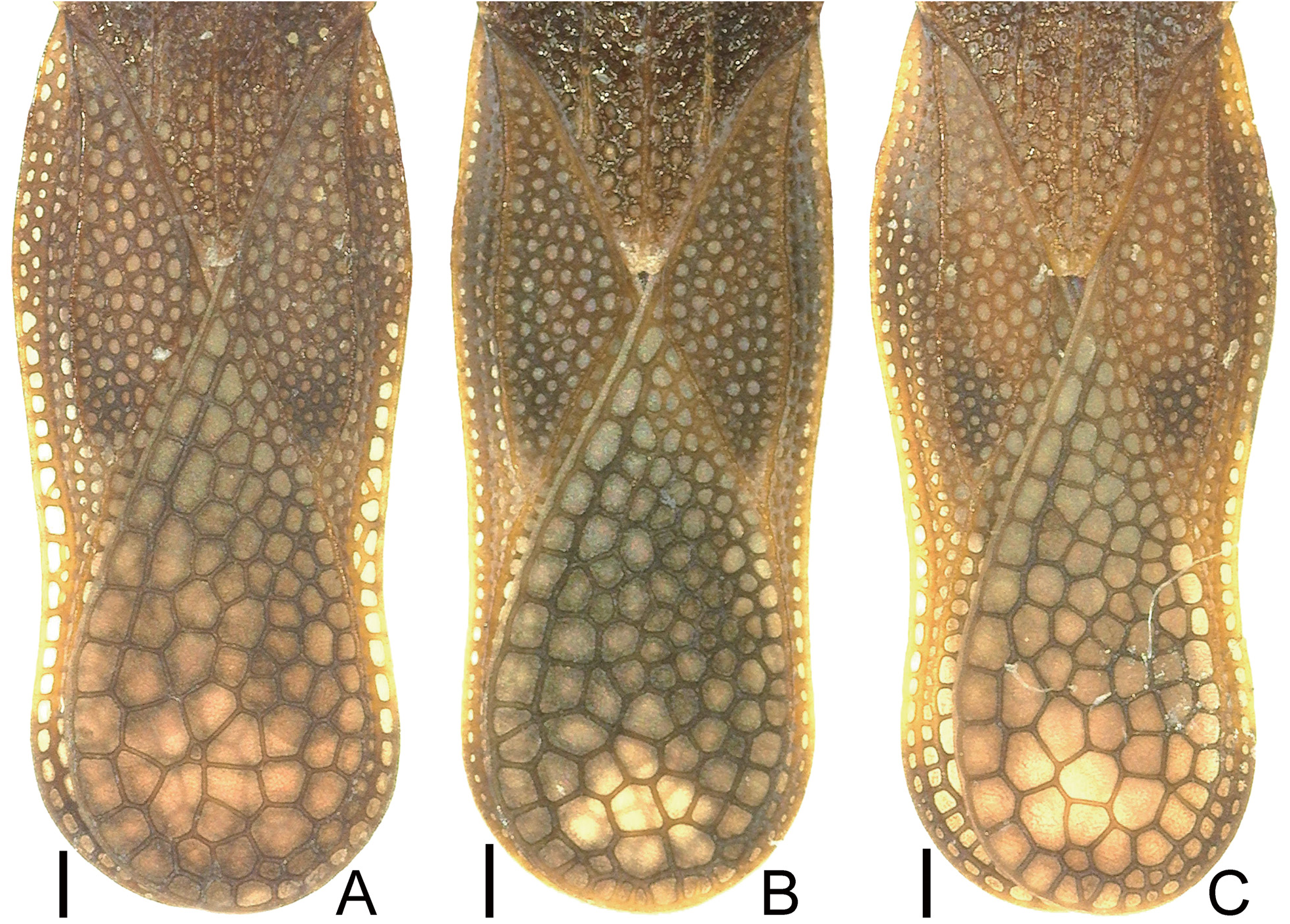
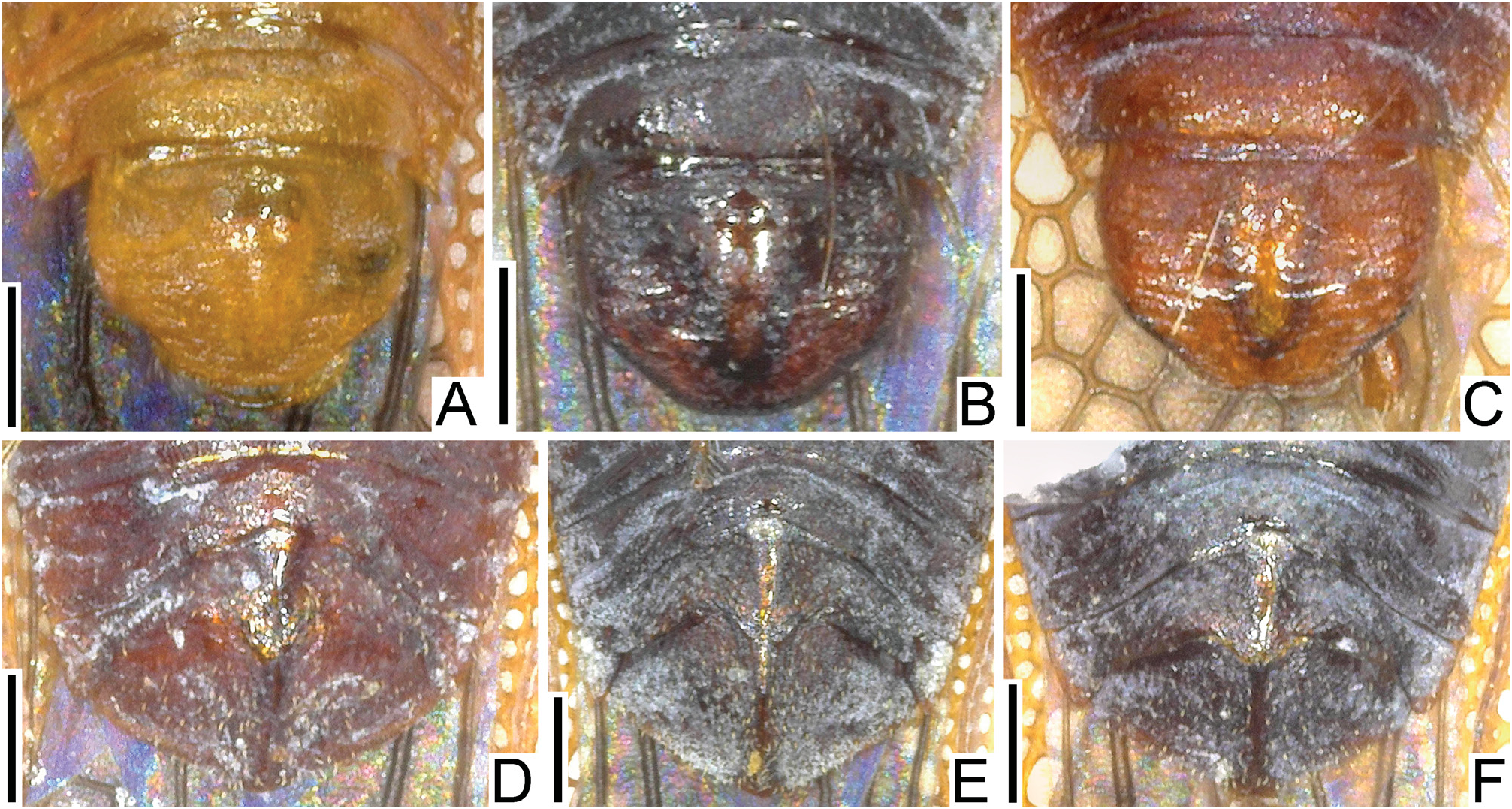
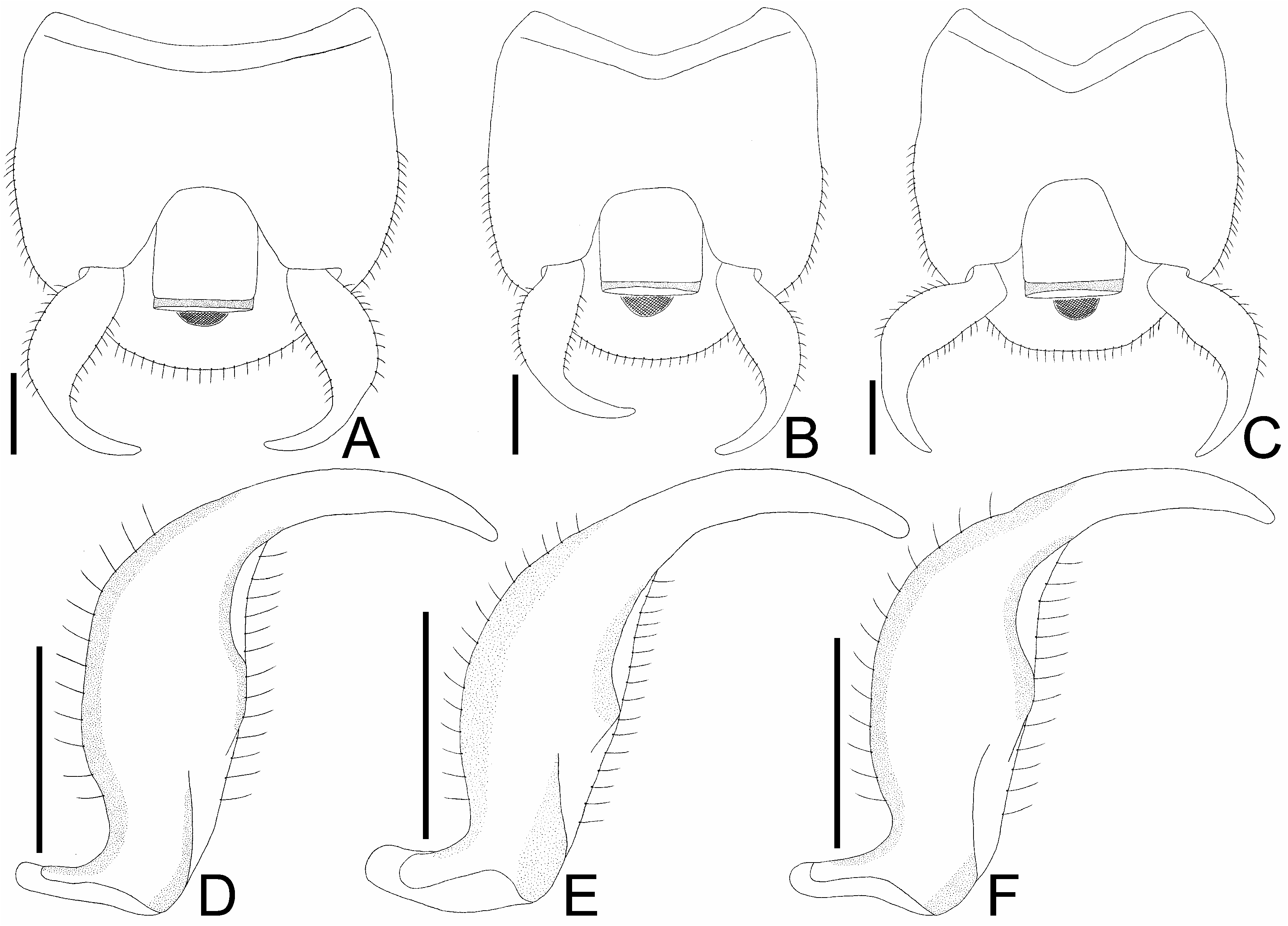
Diagnosis. Recognized among other species of Perissonemia by a combination of the following characters: body length 3.7–4.1 mm ( Figs 1A, B View Figs 1 , 2A, B View Figs 2 ); pronotal disc, posterior process and hemelytron except for areolae brown; frontal and median spines distinct ( Fig. 3A View Figs 3 ); occipital spine reaching middle part of compound eye; buccula with 3 rows of areolae at highest part; rostrum not reaching beyond posterior margin of mesosternum ( Fig. 7A View Figs 7 ); lateral carina of pronotum present on pronotal disc and posterior process ( Fig. 4A View Figs 4 ); costal area of hemelytron more than 0.5 times as wide as subcostal area at widest part of each, with 2 rows of areolae in basal part and a single row (occasionally 2 rows in very small sections) in remaining parts ( Fig. 5A View Figs 5 ); subcostal area less than 0.5 times as wide as discoidal area at widest part of each, with 3 rows of areolae at apex and 2 rows (occasionally 3 rows in very small sections) in remaining parts ( Fig. 6A View Figs 6 ); discoidal area with 6 rows of areolae at widest part; sutural area with 7 rows of areolae at widest part; and anterior margin of pygophore weakly concave in middle part of dorsum ( Fig. 9A View Figs 9 ).
Redescription. Male. Head, antennae, bucculae, pronotum, hemelytra except for most of areolae, legs and ventral surface brown; compound eye dark red; areolae of hemelytron except for sutural area translucent; pubescence on body yellowish ( Figs 1A View Figs 1 , 2A View Figs 2 , 3A View Figs 3 , 4A View Figs 4 , 5A View Figs 5 , 6A View Figs 6 , 7A View Figs 7 , 8A View Figs 8 ). Body 3.3 times as long as maximum width across hemelytra ( Fig. 1A View Figs 1 ). Head ( Figs 3A View Figs 3 , 4A View Figs 4 , 7A View Figs 7 ) covered with pubescence; pair of frontal spines distinct, touching each other at apices, reaching apex of clypeus; median spine distinct, as long as frontal spines, reaching beyond bases of frontal spines; pair of occipital spines longer than median spine, reaching middle part of compound eyes; antenniferous tubercles obtuse, slightly curved inward; clypeus smooth. Lateral margin of compound eye round. Antenna covered with pubescence; segment I cylindrical; segment II conical, shortest among antennal segments; segment III longest among antennal segments; segment IV cylindrical, longer than segment I. Buccula with 3 rows of areolae throughout its length. Rostrum not reaching beyond posterior margin of mesosternum.
Pronotum ( Figs 3A View Figs 3 , 4A View Figs 4 ) 1.6 times as long as maximum width across humeri, glabrous. Pronotal disc coarsely punctate. Hood absent. Calli smooth, partly covered with wax. Collar with 2 rows of areolae throughout its width, with anterior margin slightly curved outward. Pronotal carinae without distinct areolae. Median carina straight, extending to apex of posterior process. Lateral carina present on pronotal disc and posterior process. Posterior process of pronotum triangular, obtuse at apex.
Hemelytron ( Figs 2A View Figs 2 , 5A View Figs 5 , 6A View Figs 6 ) 2.7 times as long as its maximum width, extending beyond apex of abdomen, glabrous; maximum width across hemelytra 1.2 times as long as maximum width across humeri; costal area more than 0.5 times as wide as subcostal area at widest part of each, with 2 rows of areolae in basal part and single row (occasionally 2 rows in very small sections) in remaining parts; subcostal area less than 0.5 times as wide as discoidal area at widest part of each, with 3 rows of areolae at apex and 2 rows (occasionally 3 rows in very small sections) in remaining parts; discoidal area with 6 rows of areolae at widest part; sutural area with 7 rows of areolae at widest part.
Thoracic pleura ( Fig. 2A View Figs 2 ) coarsely punctate. Prosternum ( Fig. 7A View Figs 7 ) narrower than mesosternum. Sternal laminae lower than bucculae; anterior and posterior margins lower than lateral margin; prosternal lamina nearly straight, lower than mesosternal lamina; mesosternal lamina as high as metasternal lamina. Legs ( Fig. 1A View Figs 1 ) smooth, covered with pubescence; femora thickest in middle.
Abdomen oblong in dorsal and ventral views. Pygophore ( Figs 8A View Figs 8 , 9A View Figs 9 ) compressed dorsoventrally, hexagonal in ventral view, covered with pubescence; anterior margin weakly concave in middle part of dorsum. Paramere ( Fig. 9D View Figs 9 ) expanded in middle part, angularly curved inward in apical part; outer and inner margins covered with pubescence in middle part.
Measurements (n = 20). Body length with hemelytra 3.7–3.9 mm; maximum width across hemelytra 1.1–1.2 mm; length of antennal segments I to IV 0.3 mm, 0.1 mm, 1.2 mm, and 0.5 mm, respectively; pronotal length 1.5–1.6 mm; pronotal width across humeri 1.0 mm; hemelytral length 2.6–2.8 mm; maximum width of hemelytron 1.0– 1.1 mm.
Female. General appearance very similar to that of male ( Figs 1B View Figs 1 , 2B View Figs 2 , 8D View Figs 8 ) except for the following characters: body 3.2 times as long as maximum width across hemelytra; antennal segment III shorter than in male; maximum width across hemelytra 1.3 times as long as maximum width across humeri; terminalia pentagonal in ventral view.
Measurements (n = 20). Body length with hemelytra 3.7–4.1 mm; maximum width across hemelytra 1.2–1.4 mm; length of antennal segments I to IV 0.3 mm, 0.1 mm, 1.0– 1.1 mm, and 0.5 mm, respectively; pronotal length 1.5–1.7 mm; pronotal width across humeri 1.0– 1.1 mm; hemelytral length 2.6–2.9 mm; maximum width of hemelytron 1.0– 1.1 mm.
Remark. Although the type locality of P. occasa was merely indicated as “Japan” ( DRAKE 1942), the label on the holotype mentions “Japan: Harima” [= Japan: Honshu, southwestern part of Hyogo-ken] ( UNITED STATES NATIONAL MUSEUM OF NATURAL HISTORY 2021). The morphological species from Honshu described above match well the holotype and the original description ( DRAKE 1942) in terms of morphological characteristics. Consequently, “true” P. occasa was definitely described from Honshu.
Differential diagnosis. Among the East Asian species, Perissonemia occasa is most similar to P. gressitti in the morphological characteristics. However, based on a comparison between a number of non-type materials together with the photograph of the holotype of P. occasa ( UNITED STATES NATIONAL MUSEUM OF NATURAL HISTORY 2021) and the original description ( DRAKE & POOR 1936) together with the photographs of the holotype of P. gressitti ( UNITED STATES NATIONAL MUSEUM OF NATURAL HISTORY 2021), two main characteristics were recognized to differentiate P. occasa from P. gressitti ; discoidal area of hemelytron with 6 rows of areolae at widest part ( Fig. 5A View Figs 5 ) (5 rows in P. gressitti ); and sutural area with 7 rows of areolae at widest part (6 rows in P. gressitti ). Morphological differences between this species and the other two Japanese species are provided in the identification key below.
Host plant. Osmanthus heterophyllus , “Hiiragi” ( Oleaceae ) ( Fig. 10C View Figs 10 ) ( NAKANO 1984, TSUKADA 1995, YAMADA & TOMOKUNI 2012; present study). Perissonemia occasa feeds only on this oleaceous plant in the field and is monophagous, similarly to many tingids ( SCHUH & WEIRAUCH 2020). However, this lace bug sometimes occurs on planted O. insularis within its distribution range ( TSUKADA 1995) and feeds on O. × fortunei (a hybrid of O. fragrans and O. heterophyllus ) and O. fragrans var. aurantiacus in captivity ( NAKANO 1984).
Biology. Perissonemia occasa feeds on the abaxial surface of the leaves of the aforementioned host plant ( TSUKADA 1995; present study), similarly to many tingids ( SCHUH & WEIRAUCH 2020). This lace bug is considered to be univoltine ( TSUKADA 1995); adults were collected in almost all seasons ( DRAKE 1942, TAKEYA 1962, NAKANO 1984, TSUKADA 1995; present study); nymphs were observed in June and July ( TSUKADA 1995); and the overwintering stage appears to be adult because of the holotype collected in March (cf. DRAKE 1942).
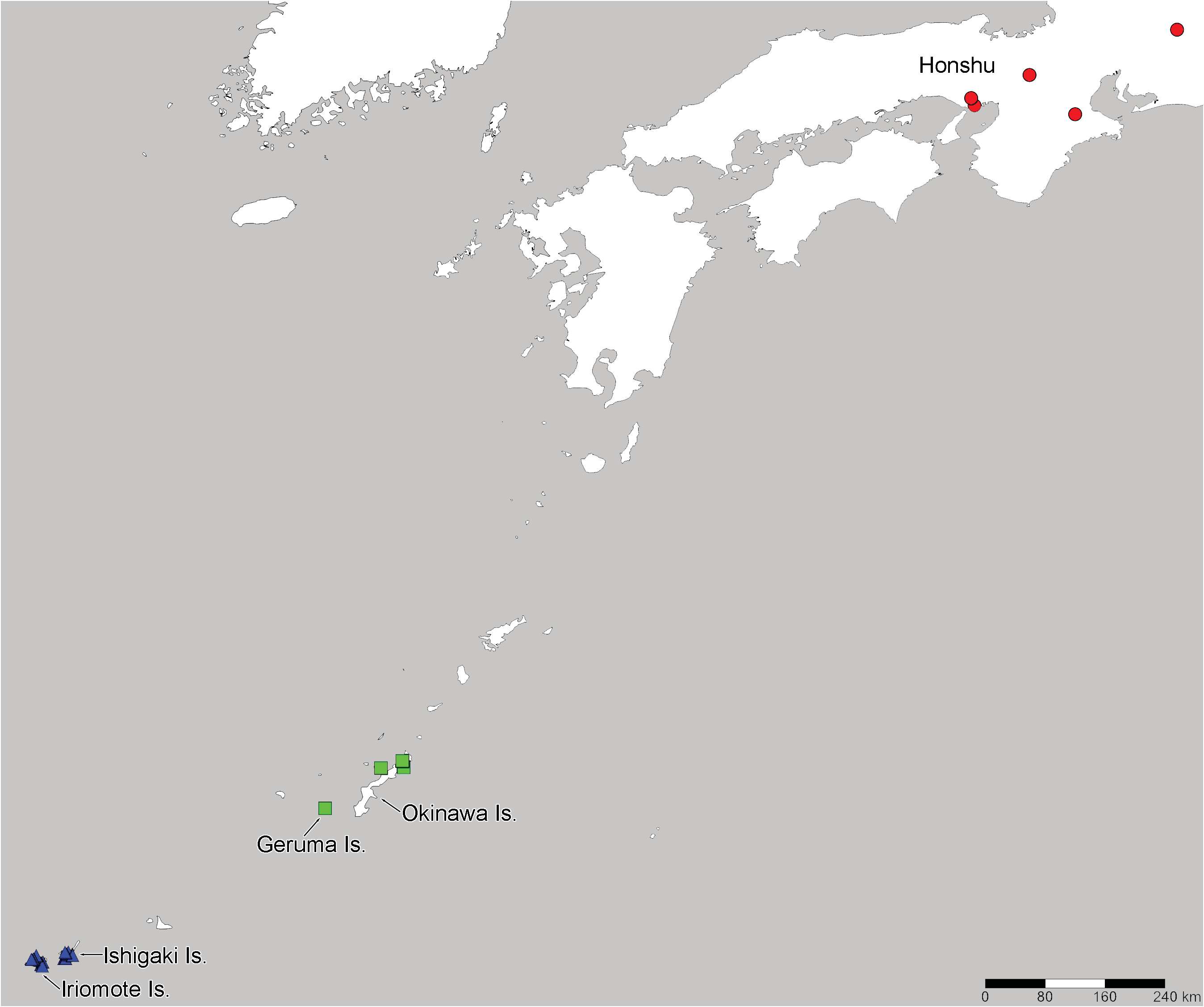
Distribution. Japan (Honshu) ( Fig. 11 View Fig ) ( DRAKE 1942; TAKEYA 1962; NAKANO 1984; TSUKADA 1995).
Previous records from China ( DRAKE & RUHOFF 1961, JING 1981) do not list the examined specimens and appear to be erroneous. Perissonemia occasa inhabits laurilignosa in the temperate climate of Honshu, which is located in the Palaearctic Region.







| NSMT |
National Science Museum (Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Perissonemia occasa Drake, 1942
| Souma, Jun 2022 |
Baeochila occasa: LEE (1969)
| LEE C. E. 1969: 233 |
Perissonemia occasa: TAKEYA (1951)
| YAMADA K. & ISHIKAWA T. 2016: 432 |
| YAMADA K. & TOMOKUNI M. 2012: 198 |
| TSUKADA M. 1995: 49 |
| TAKAHASHI T. 1990: 3 |
| MIYAMOTO S. & YASUNAGA T. 1989: 167 |
| NAKANO K. 1984: 482 |
| JING H. - L. 1981: 311 |
| DRAKE C. J. & RUHOFF F. A. 1965: 324 |
| TAKEYA C. 1962: 56 |
| DRAKE C. J. & RUHOFF F. A. 1961: 135 |
| TAKEYA C. 1951: 18 |
Perissonemia occasa
| DRAKE C. J. 1942: 2 |