
Adineta beysunae, Örstan, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4524.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:6670AAB9-C770-4B62-A462-090C680FBB46 |
|
DOI |
https://doi.org/10.5281/zenodo.5959248 |
|
persistent identifier |
https://treatment.plazi.org/id/8029878D-FFB1-E009-FF24-AEE5A5514150 |
|
treatment provided by |
Plazi |
|
scientific name |
Adineta beysunae |
| status |
sp. nov. |
Adineta beysunae sp. nov.
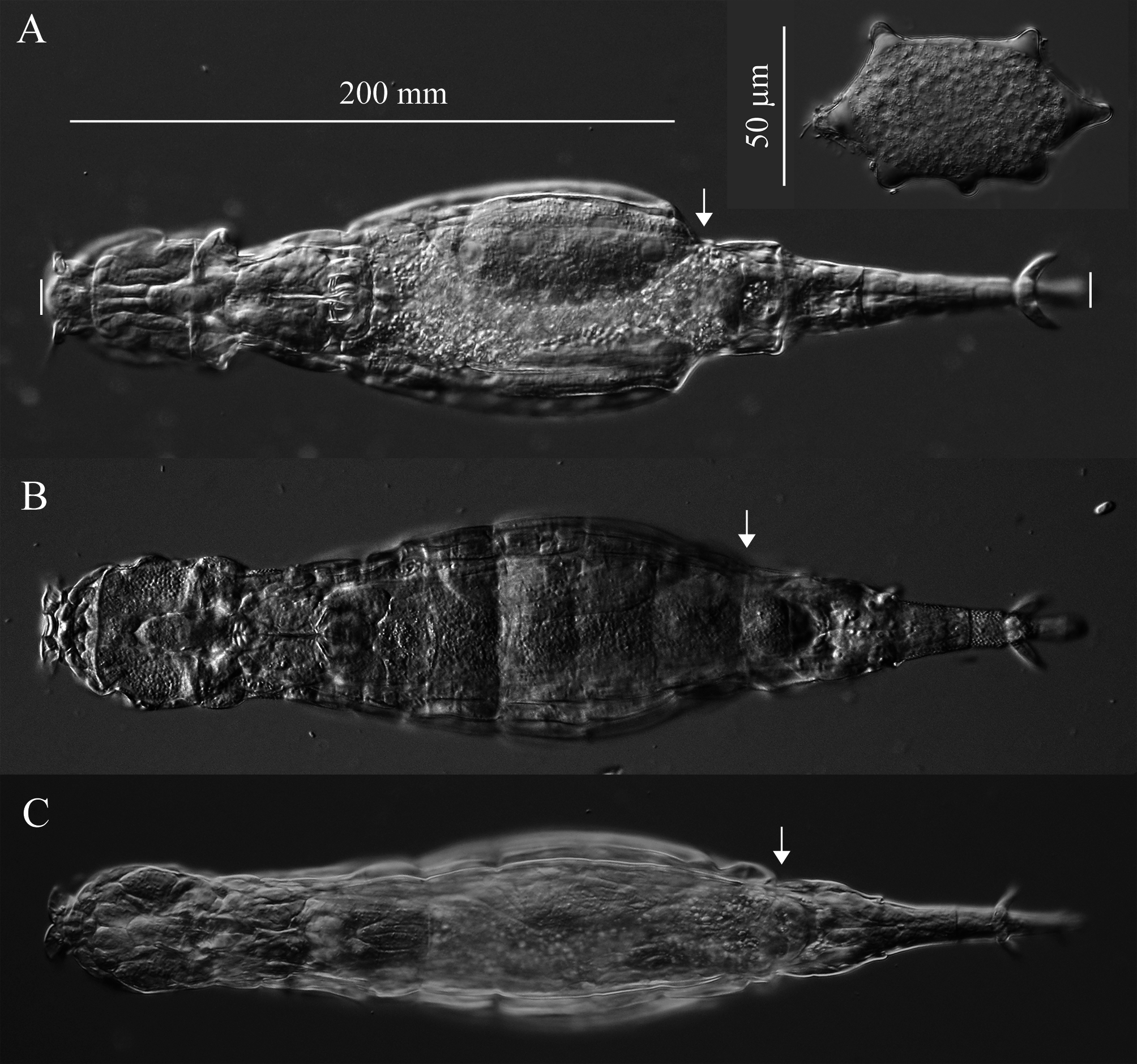
( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 5A View FIGURE 5 , 6 View FIGURE 6 , 7 View FIGURE 7 , 8 View FIGURE 8 ; Table 2)
Material. The description of A. beysunae was based on 25 specimens constituting the type series. The holotype, stained with neutral red and mounted in glycerine jelly between two cover glasses and secured in a Cobb’s slide holder ( Fig. 4A, B View FIGURE 4 ), has been deposited in the Academy of Natural Sciences, Philadelphia, PA, U.S.A. (ANSP 2126). Seven paratypes are in the collection of the author identified with the private variant designation b1.P 170623. All photographs in this description are those of the type specimens.
Type locality. The garden of a private residence, Germantown, Maryland, USA (39.2075 N, 77.2554 W). The microhabitat of the species was a container filled with rain water and plant debris GoogleMaps .
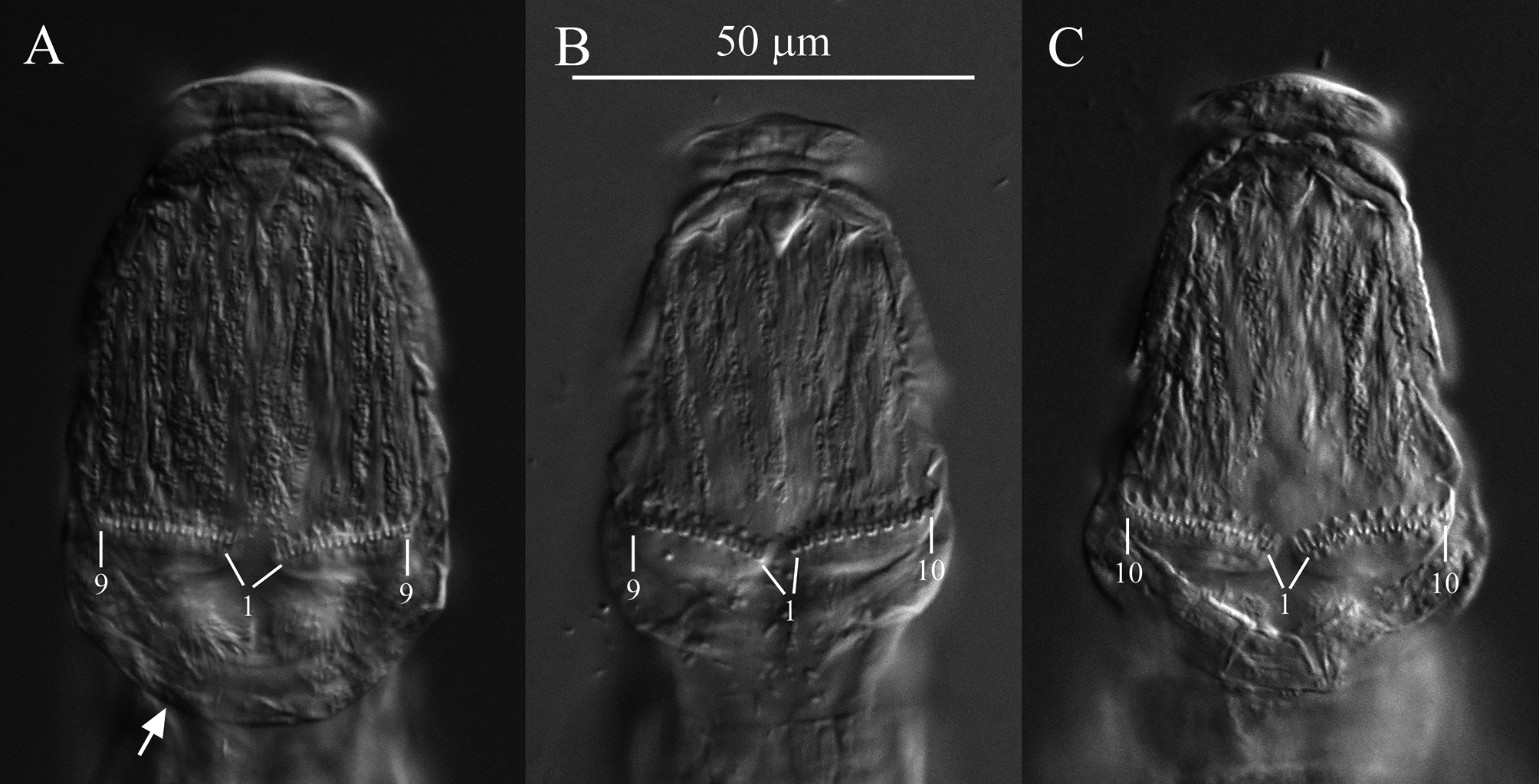
Diagnosis. Each rake with 9 to 10 U-gaps ( Fig. 5 View FIGURE 5 ). Posterior end of first rump pseudosegment with lateral swellings; first and second foot pseudosegments each with a dorsal thickening appearing as peg-like lateral projections ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 8A View FIGURE 8 ).
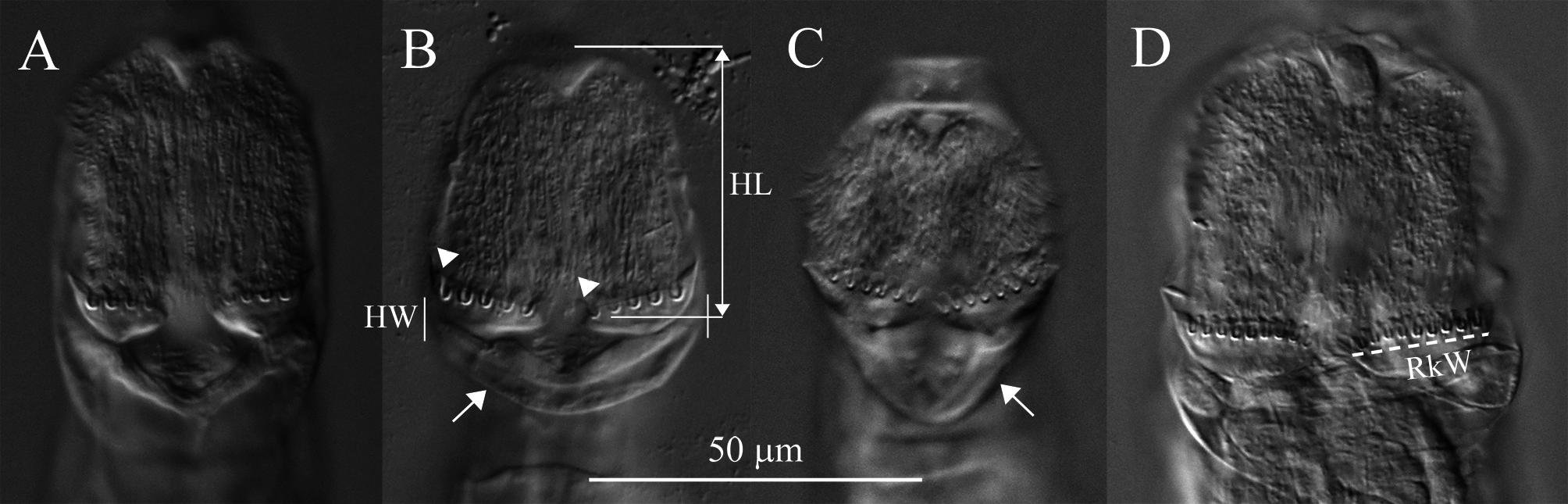
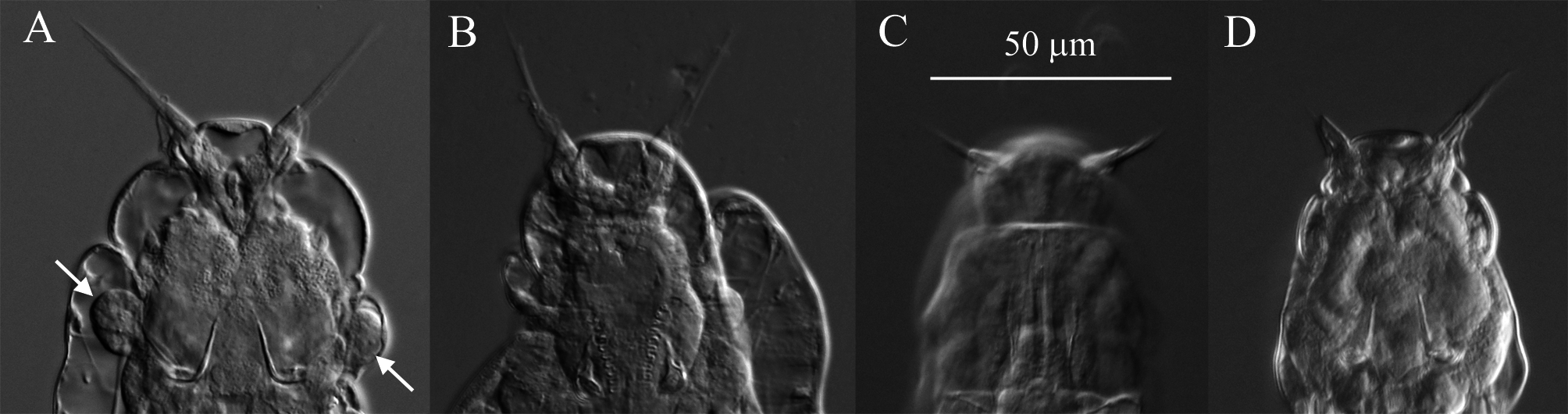
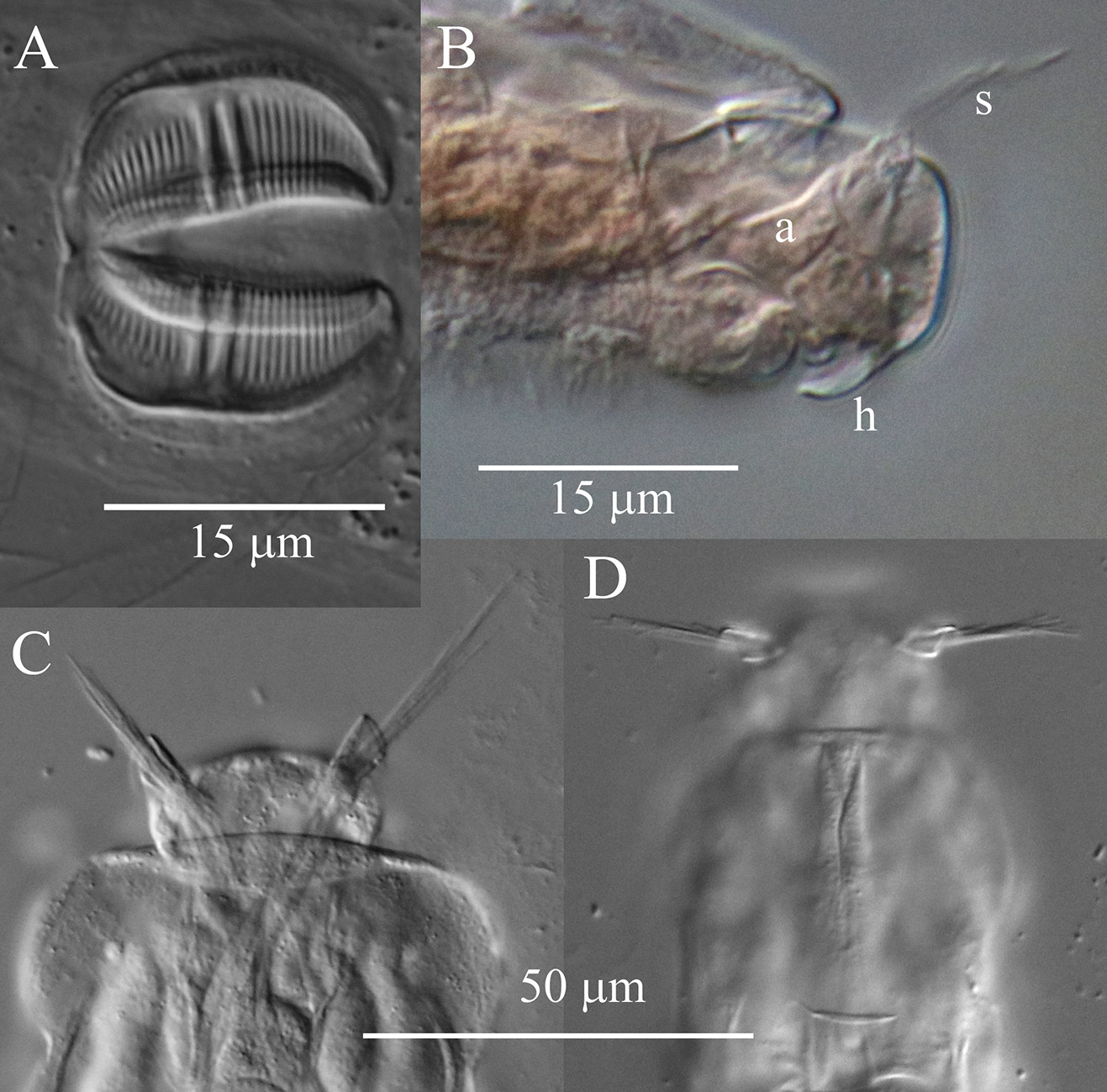
Description. Integument on dorsal rostrum folds into anterolateral auricles through which bundles of setae emerge ( Figs. 6 View FIGURE 6 , 7B, C View FIGURE 7 ). Each bundle with probably at least four setae of often unequal and variable lengths ( Figs. 6 View FIGURE 6 , 7C View FIGURE 7 ); longest measured seta: ~30 µm. Ventral rostrum with short cilia. Length of antenna same or slightly less than half lateral width of neck where antenna attaches. Integument lateral to rake organ juts out accommodating a gland-like organ ( Figs. 4 View FIGURE 4 , 6A View FIGURE 6 ). Lower lip broad, uniformly curved ( Fig. 5 View FIGURE 5 ). Numbers of U-gaps on rakes ( Fig. 5 View FIGURE 5 ): 9– 9 (n = 3), 9–10 (n = 3), 10–10 (n = 8, including holotype). RkW (n = 7; all with 10–10 U-gaps): 20.1–22.4 µm; mean: 21.4 µm. Dental formula of trophi, 2/2 ( Fig. 7A View FIGURE 7 ). Trophi length (n = 7): 17.0–18.6 µm; mean: 17.6 µm.
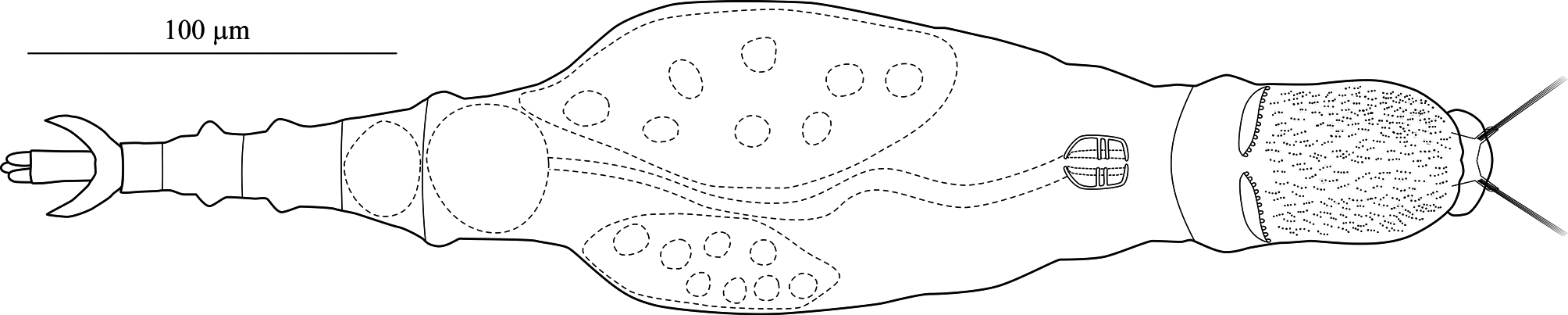
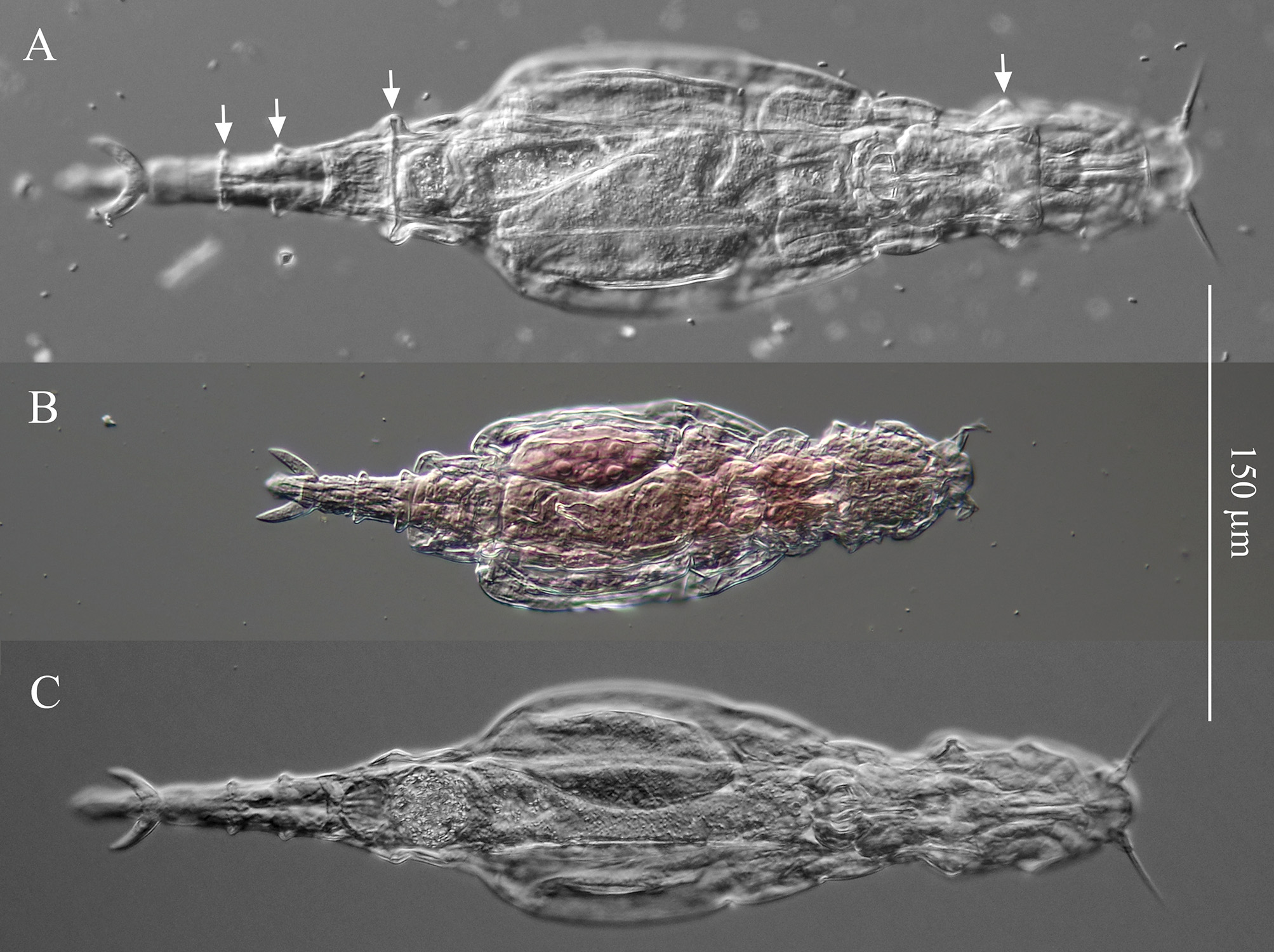
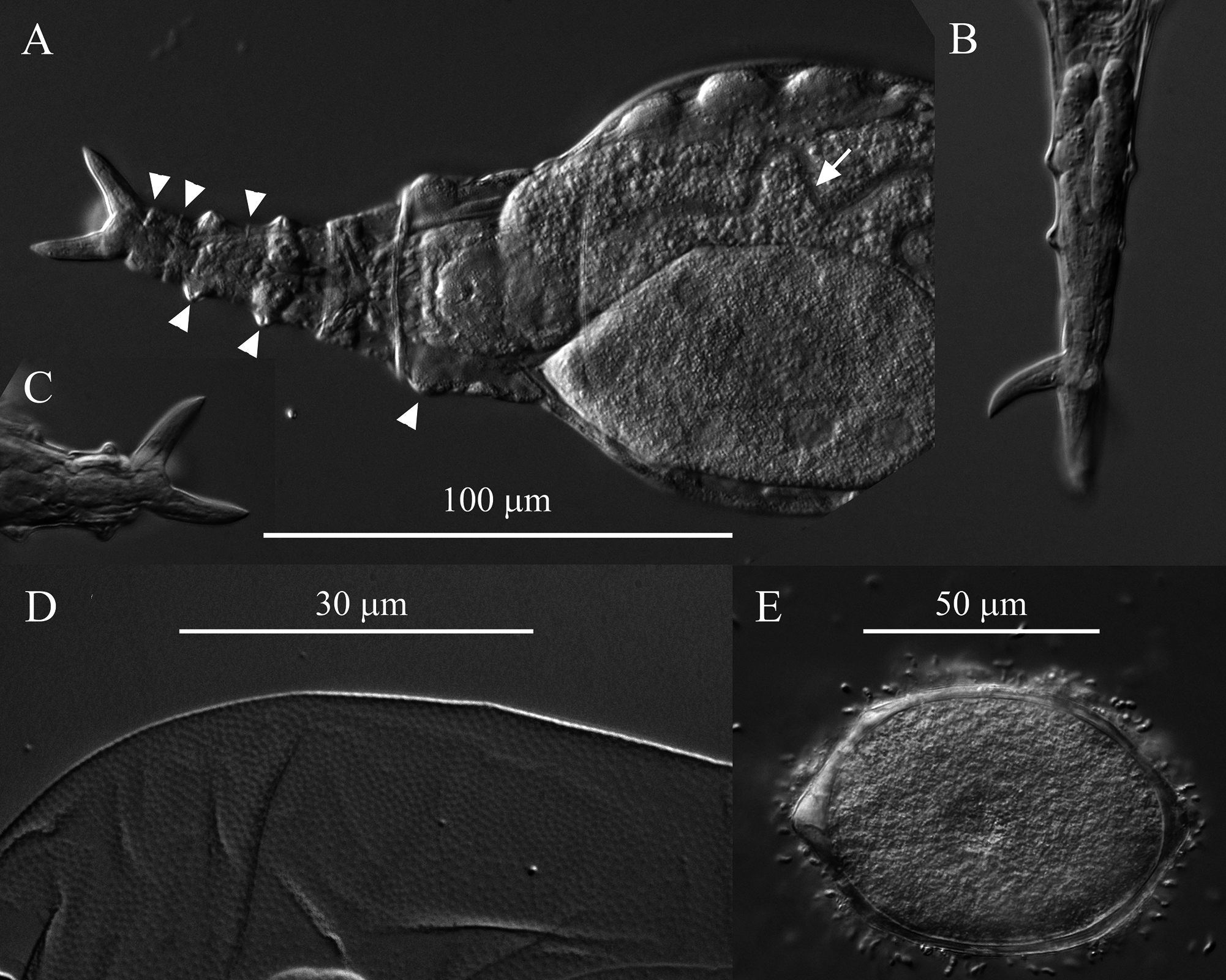
Stomach with lumen ( Fig. 8A View FIGURE 8 ). Each vitellarium with 8 nuclei. Posterior end of central trunk narrows somewhat steeply into rump ( Fig. 4 View FIGURE 4 , 8A View FIGURE 8 ). Lateral corners of posterior end of first rump pseudosegment bulging ( Figs. 4 View FIGURE 4 , 8A View FIGURE 8 ); second rump pseudosegment passes into foot smoothly. Foot with 3 pseudosegments prior to spurs; demarcation between first and second pseudosegments indistinct ( Fig. 8A View FIGURE 8 ). First and second foot pseudosegments with narrow dorsal thickenings forming blunt lateral projections ( Figs. 4 View FIGURE 4 , 8A View FIGURE 8 ). Spurs blade-like with short straight interspace between bases ( Figs. 4 View FIGURE 4 , 8 View FIGURE 8 ). Spur length (n = 14): 16–22 µm. Three toes. Integument all over body finely and densely stippled most distinctly on trunk and first rump pseudosegment ( Fig. 8D View FIGURE 8 ). Body without noticeable color. TL (n = 20; undetermined ages): 310–412 µm; three longest specimens: 397 µm, 406 µm (holotype) and 412 µm. Mean RF /TL (n = 10): 0.38; mean HL/HW (n = 6): 1.1. Egg elongated hexagonal with nipple-like protuberance at each pole ( Fig. 8E View FIGURE 8 ). Mean egg dimensions (n = 8): 84 x 53 µm. Additional dimensional data in Table 2. a Absolute dimensions are in µm. TL: total length; TrL: truncated length; HL/HW: ratio of head length (HL) to head width (HW); TL/BW: ratio of total length (TL) to body width (BW); SL: spur length; TiL: trophi length; RkW: rake width; EL: egg length; EW: egg width.
Etymology. The species is named after my wife Beysun Örstan.
Remarks on morphology. Unobstructed A. beysunae creeps fast with the rostral setae and the spurs curving up; while the setae point ahead of the rostrum, the spurs are directed posteriorly ( Figs. 4 View FIGURE 4 , 8B View FIGURE 8 ). Active animals were difficult to examine at high magnifications unless they were slowed down by the removal of water from under the cover glass. The variability of the lengths of the setae of A. beysunae ( Fig. 6 View FIGURE 6 ) indicates that the setae length of a variant may not always be a reliable diagnostic trait. I have not determined if the setae lengths change because they are retractable or because they are sometimes dropped and then regrown. The anterolateral auricles on the rostrum are folds of the integument in the shape of tube-like sleeves that are open along their lateral edges ( Figs. 6A View FIGURE 6 , 7B, C View FIGURE 7 ). The integument on the ventral surface of the rostrum forms a free edge (lamella) that laterally appears as a “hook” ( Fig. 7B View FIGURE 7 ). This hook is not diagnostic, because a similar morphology is present in many Adineta variants (for example, Davis 1873). The short cilia present on the ventral side of the rostrum are most easily seen on preserved specimens using oil immersion. The flexibility of the head causes its shape to vary as a specimen moves and also among specimens ( Fig. 5 View FIGURE 5 ). One trophi that was 14.1 µm long was excluded from the sample used to calculate the mean given above. The insides of the lumen and the intestine are ciliated. The lateral projections on the first and the second foot pseudosegments are the continuations of the dorsal thickening present at about the middle of each pseudosegment ( Figs. 4 View FIGURE 4 , 8A View FIGURE 8 ). The lateral outline of a spur curves more or less uniformly on the outside, but widens near the base on the inside; the overall spur shape is best explained with photographs ( Fig. 8B, C View FIGURE 8 ). The stippled appearance of the integument of A. beysunae is caused by very small circular formations that are most easily seen on compressed live animals or on specimens whose insides have been cleared with SDS ( Fig. 8D View FIGURE 8 ).
Comparison with other species. The diagnostic traits of A. beysunae form a unique combination absent in any of the described Adineta species ( Donner 1965; Iakovenko et al. 2015; Örstan & Plewka 2017). The general shape of the rostrum with long setae, the outlines of the posterior trunk and the spurs of A. beysunae do resemble those of A. steineri Bartoš 1951 ( Steiner 1913; Donner 1965). The latter was described by Steiner (1913) as A. barbata nov. var.? and erected as a distinct species by Bartoš (1951) without additional morphological information. A. steineri is one of those species that have incomplete original descriptions; it is not possible to match its original description conclusively with a known variant.
Steiner (1913) did not give any dimensions or include a scale with his drawings. Moreover, as Wulfert (1960) already noted, the proportions of the body parts of the rotifer in Steiner’s drawing look odd: the head is too big relative to the trunk, while the foot is too short and narrow. Steiner could not state definitely the number of rake denticles of his specimens, but his drawing shows 10 on each rake, which would equal 9 U-gaps. Wulfert (1960) did not mention the rake organ in his brief description of A. steineri , but his drawing seems to show only five denticles on each rake, although this was possibly not meant to be a realistic rendition of the rakes. Donner (1965) did not mention the number of denticles of A. steineri either, but his drawing shows 8 denticles (7 U-gaps) on each rake. Haigh (1968) gave a long description of his specimens of A. steineri , but neither mentioned the rake organ nor included it in his drawing. Song & Kim (1996) wrote that A. steineri had “8 sharp claws” on its rake and showed as many in their drawing. These literature discrepancies in the numbers of rake denticles of A. steineri may have resulted from either careless descriptions or the presence of several variants, lumped under the name A. Steineri , that differ at least in their rakes. There is also some uncertainty concerning the foot morphology of the specimens identified as A. steineri . The feet of the rotifers in the drawings of Steiner (1913) and Donner (1965) had no protuberances. Wulfert’s (1960) drawing of A. steineri shows the two pre-spur pseudosegments of the foot with slight lateral swellings (not mentioned in the text), while Haigh (1968) did note that the foot of his specimen had “two characteristic swellings” and showed these in his drawing. However, these swellings do not resemble the distinct protuberances present on the foot of A. beysunae . An additional uncertainty associated with the identification of A. steineri is the morphology of its egg. Steiner (1913) did not mention the egg of his specimens. Donner (1965) and Haigh (1968) described the egg of A. steineri with 7 and 6 protuberances, respectively, but Voigt (1957) claimed the species was viviparous.
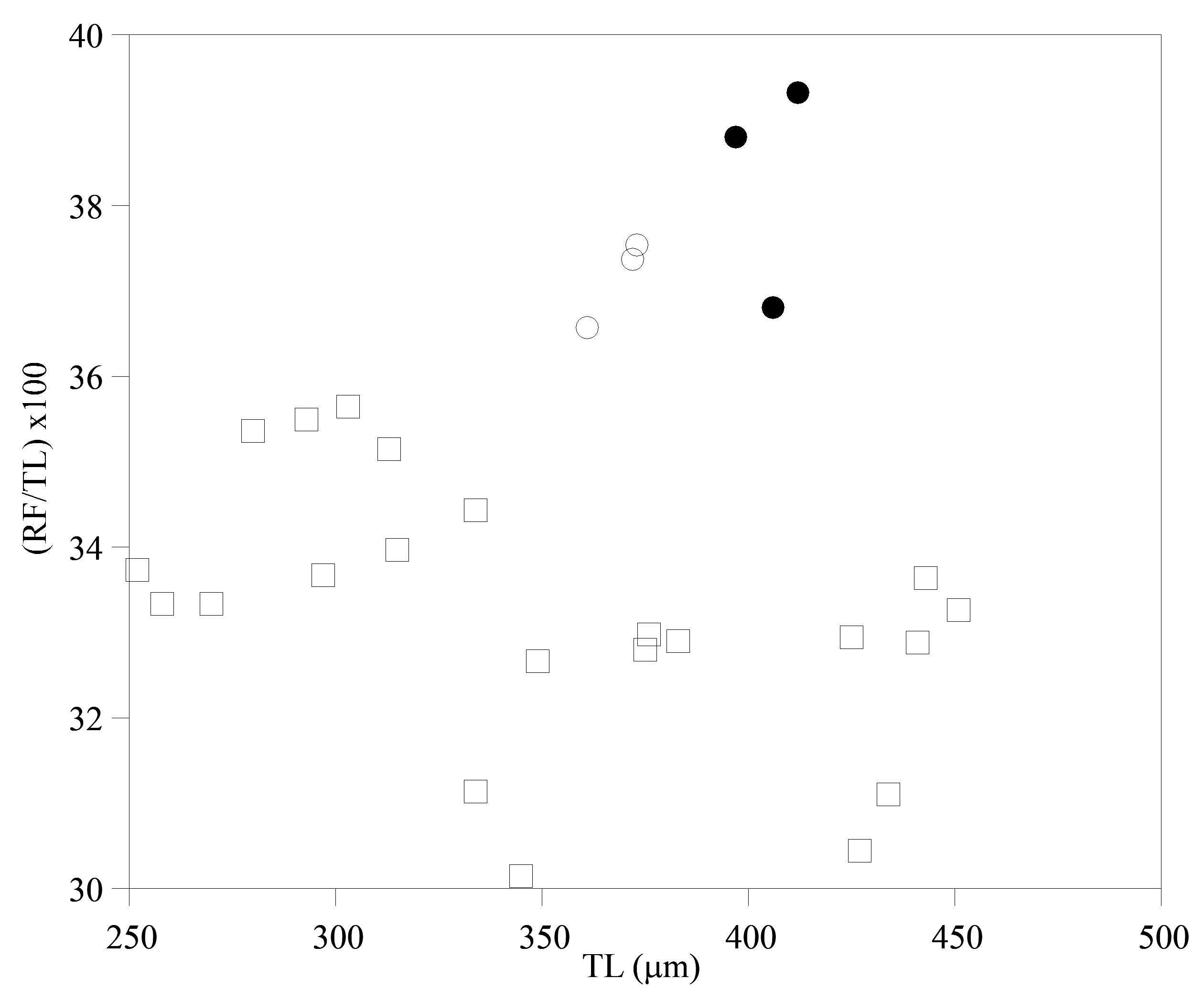
I am tentatively identifying as A. cf. steineri a variant from a sample of roof moss from England ( Figs. 1A View FIGURE 1 , 2D View FIGURE 2 , 6D View FIGURE 6 ). Adineta beysunae and A. cf. steineri have the largest relative RF lengths among the Adineta variants I have examined ( Fig. 9 View FIGURE 9 ). However, in A. cf. steineri the lateral corners of the posterior end of the first rump pseudosegment were not as prominent as those of A. beysunae and the foot did not have the thickenings present in the latter species ( Fig. 1A View FIGURE 1 ). Moreover, the rakes of 3 specimens of A. cf. steineri examined all had 9–9 U-gaps ( Fig. 2D View FIGURE 2 ). Also, the egg of A. cf. steineri , with 7 nipple-like protuberances ( Fig. 1A View FIGURE 1 ), was unlike the egg of A. beysunae ( Fig. 8E View FIGURE 8 ). Regarding the rostral setae of A. steineri, Steiner (1913) , Haigh (1968) and Song & Kim (1996) stated that there was only one seta (bristle) on each side, while Donner (1965) in an unclear remark thought that each bristle was composed of several setae. I observed bundles of setae protruding from the auricles of A. cf. steineri ( Fig. 7D View FIGURE 7 ). In summary, despite some similarities in their morphologies as well as body proportions, A. beysunae and A. cf. steineri are clearly different variants.
Remarks on habitat and occurrence. Adineta beysunae was found in a container of rain water and plant debris in the garden of my house in Germantown, Maryland, USA. Two cleaned plastic containers (diameter, ~ 12 cm; depth, ~ 6 cm) were placed empty ~ 3 m apart in the garden at the start of a colonization experiment on 23 April 2017. During the following two months a grassy shrub ( Miscanthus sp.) grew around and above each container. The containers received only rain water, initially directly (when the shrubs were short) and later through the shrubs. A sample of water and plant debris was removed from each container for the first time on 23 June 2017 and specimens of A. beysunae were found in one sample but not in the other. Both samples also contained other bdelloids as well as tardigrades, nematodes, ciliates and arthropods. These organisms may have been brought into the containers by rain water or plant material falling in them or by local animals (rabbits, squirrels, snakes, frogs, slugs, etc.) that may have entered the containers or drank from them. The colonization experiment ended on 1 October 2017 when the source container of A. beysunae was found sideways and empty. On 11 November 2017 a sample of plant debris and soil was removed from the location where the source container of A. beysunae had stood and placed in water. Bdelloids appeared in this culture, but none was A. beysunae . All of the specimens in the type series of A. beysunae were from the sample of 23 June 2017. As of October 2018, small numbers of A. beysunae are being kept in laboratory cultures. However, I have not been able to optimize the culture conditions and the longterm longevity of the present cultures is uncertain. Specimens have also been saved in ethanol and in glycerol for future extraction of DNA and trophi, respectively.
The appearance of A. beysunae in only one of two containers a few meters apart and its apparent absence in the nearby soil underscores the generalization that the distributions of many bdelloids, especially the limnoterrestrial species that live in mosses and similar microhabitats, are often spatially and temporally random and unpredictable over the habitat space. Murray’s (1908) observations regarding his irreproducible collection of a bdelloid species ( Mniobia circinata ) he was describing are worth citing here: “This species affords another of those puzzles in distribution which are especially familiar to students of the Rotifera. The moss on the wall was growing in little detached cushions. One of these was casually pulled in passing, and was washed on reaching home. When the strange species was found so abundant in it, the wall was visited again, and surrounding tufts pulled, but none of the Rotifers [sic] found.” Their seemingly itinerant lifestyles and the generally impermanent nature of their microhabitats make it unrealistic to expect to find many bdelloids at their type localities or any other locality where they were once collected after the passage of even a short period. The discovery of a new bdelloid species in as mundane a habitat as a garden container also highlights the unappreciated possibility that more species may be awaiting discovery in other deceptively familiar places.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |