
Stenus canaliculatus, Gyllenhal, 1827
|
publication ID |
https://doi.org/ 10.5852/ejt.2012.13 |
|
DOI |
https://doi.org/10.5281/zenodo.3858638 |
|
persistent identifier |
https://treatment.plazi.org/id/803F87FD-C02F-FFCA-FD89-A96FFB6B9A10 |
|
treatment provided by |
Carolina |
|
scientific name |
Stenus canaliculatus |
| status |
|
Redefinition of the canaliculatus View in CoL group
Diagnosis
Body size moderate; body length of the known species: 2.8 to 4.7 mm.
Head fairly small to fairly broad in comparison to elytra and pronotum. Upper surface between eyes feebly evenly convex, nearly flat, to slightly concave as a whole; with longitudinal elevations and impressions nearly absent to well developed. Antennae rather short to long, with the club segments more or less elongated. Internal tooth of each mandible placed a little before the middle, directed only slightly dorsally of the main plane. Maxillar palpi with yellow and not shortened basal segment. A considerable reduction of the adhesion capture apparatus may be supposed for all the members of the group (for more details see the Remarks section below).
Pronotum with long and sharp longitudinal median furrow more or less deep, less often very feeble, stroke-shaped.
Elytra nearly rectangular, large, with short rounded humeri, to shortened, slightly trapezoid with humeri less developed.
Legs moderately long; the segment 4 of tarsi without emargination; segment 1 of metatarsi about the same length as to a bit longer than segment 5 but distinctly shorter than segments 2 to 4 together.
Abdomen moderately convex, with paratergites evident on abdominal segments 3 to 7; four anterior visible tergites each with four short but evident longitudinal keels at basal part; posterior margin of the tergite 7 with well-developed fine membranous fringe.
Puncturation of pronotum and elytra diverse, but always non-rugose.
Ground sculpture cellular or net-shaped to entirely absent.
Pubescence moderate, contiguous to fairly long, outstanding.
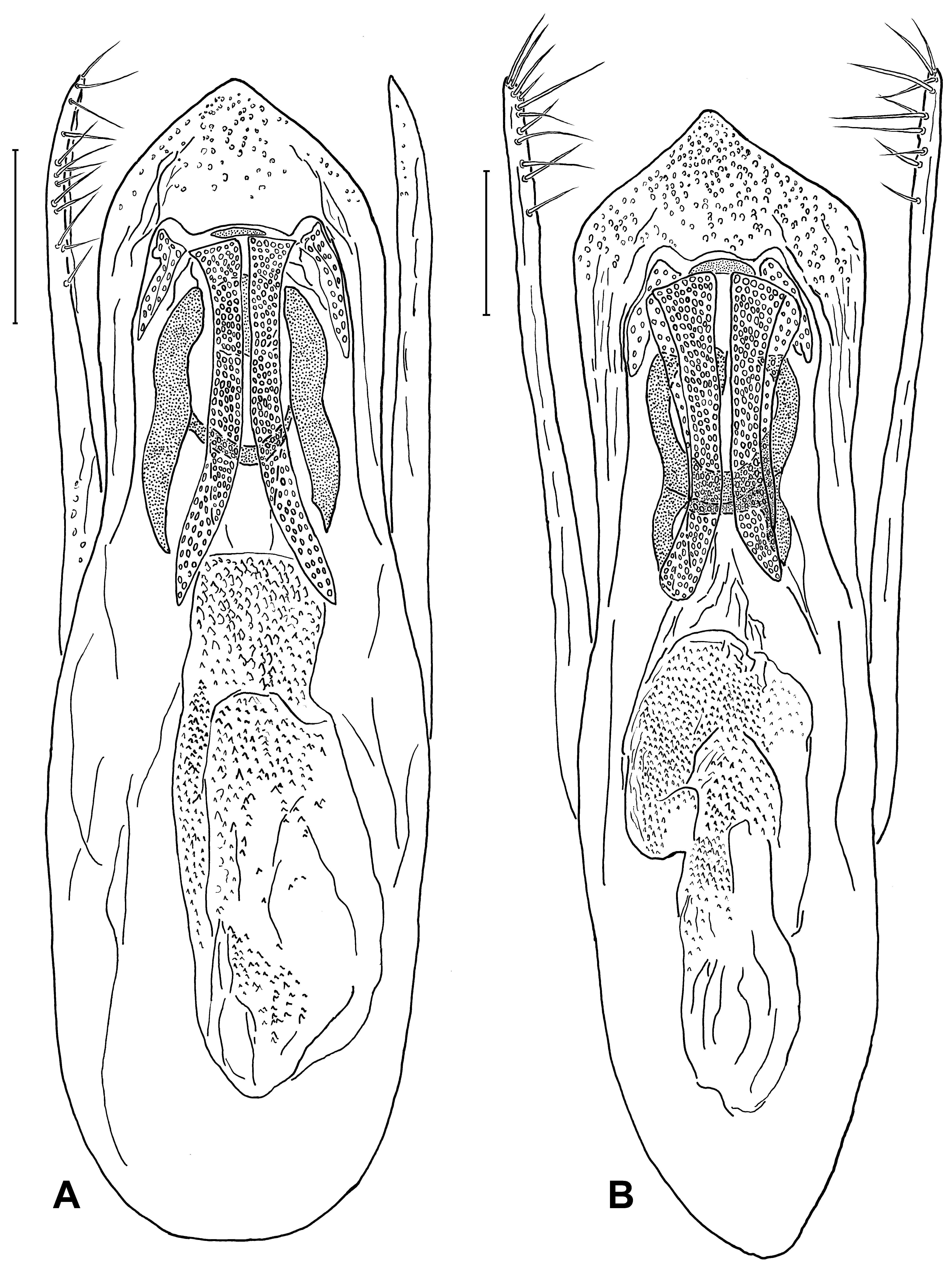
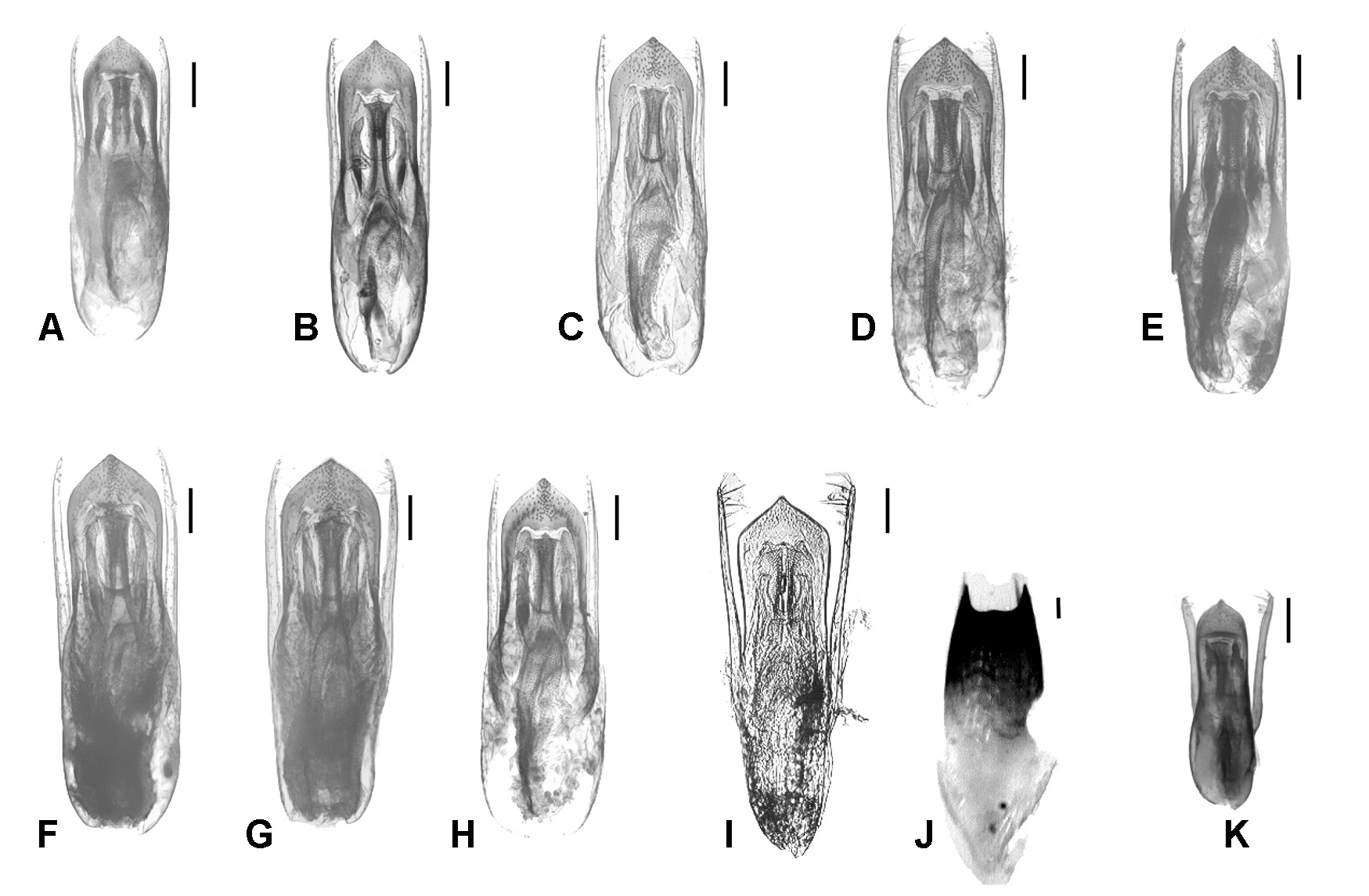
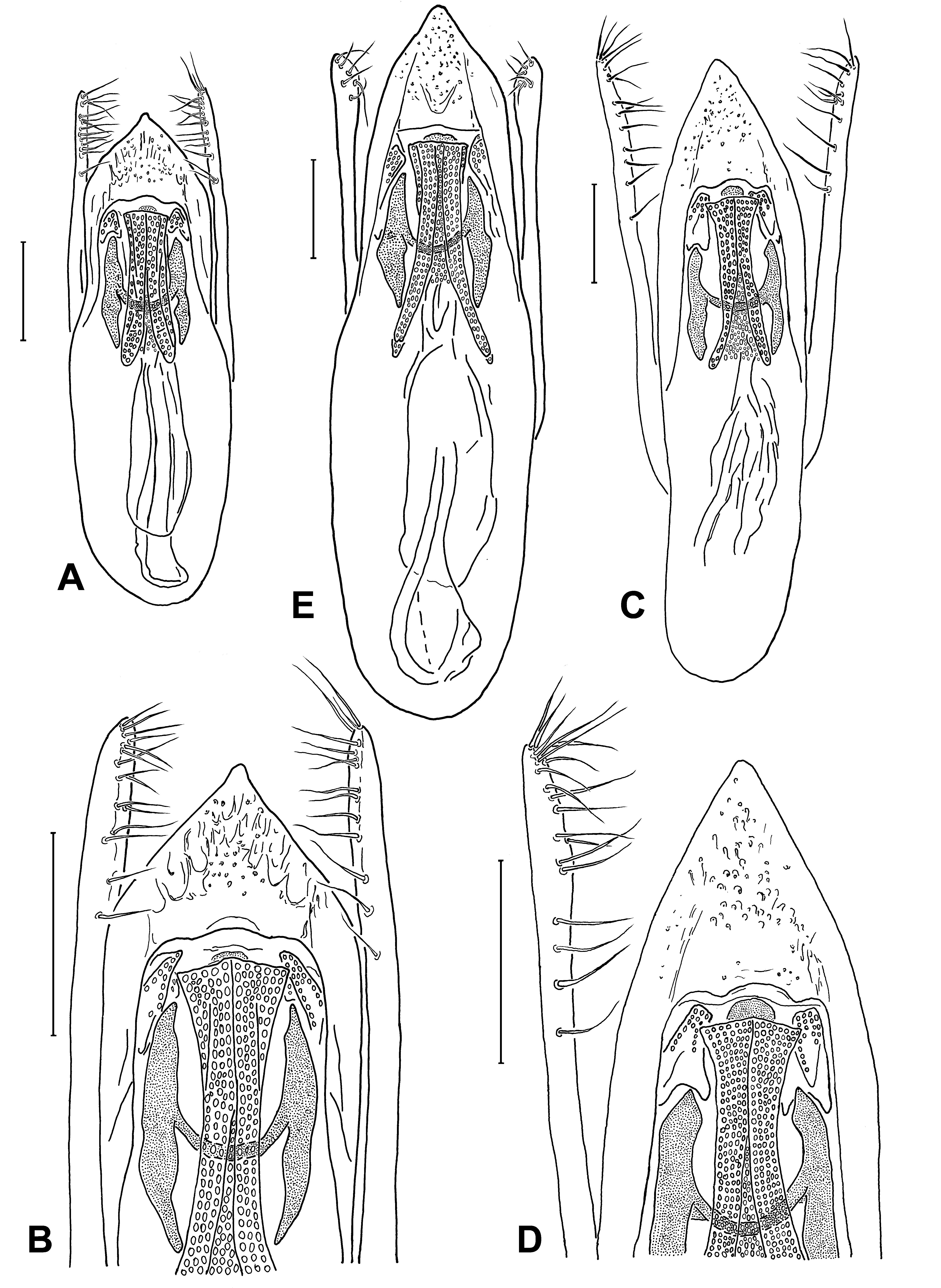
MALE. Legs without specific features; posterior margin of abdominal sternites 6–7 shallowly emarginated to about straight;abdominal sternite 8with a broad but not deep, rounded or angularly rounded emargination of posterior margin; abdominal sternite 9 with large posterolateral teeth not incurved inwards; ventral depressions of sternites 6–8, if present, flat and feeble, never flanked by sharp keels. Aedeagus relatively large; median lobe archetypically lanceolate, with apical sclerotized part to a variable extent angular to angularly rounded; endophallus with paired medial bands (optionally, an unpaired bar may also be distinct), small but visible lateral parts, and H-shaped expulsion clasp producing lateral portions fairly broad; basal tube of diverse structure, more or less sclerotized. Parameres cylindrical, with apical broadening, if present, very feeble, rather vague; apico-internal setiferous surface differentiated poorly to moderately, setae fairly uniform.
FEMALE. Posterior margin of abdominal sternite 8 broadly rounded to angularly rounded; each valvifer with posterolateral tooth directed backwards. Spermatheca sclerotized to a greater or lesser extent.
In the structure of abdomen, the male genitalia, the abdominal sternite 9 of both males and females (valvifera), the group under consideration is most closely related to the boops and palposus groups of the same subgenus Nestus . It differs from these by the structure of the mandibles, by the adhesion capture apparatus more or less shortened, by the pronotum with long and sharp, line-shaped, longitudinal median furrow, by the evidently narrower aedeagus, by the endophallus with the expulsion clasp’s transversal bridge adjoining to the lateral pieces near their middle, by the abdominal sternite 9 with posterolateral teeth not being curved inwards, by the male legs without peculiar features (not all the species of the named groups have such features well developed); from the palposus group it can be easily distinguished by the yellow basal segment of the maxillar palpi. The differences from S. (N.) melanopus Marsham, 1802 , similar externally to some species of the canaliculatus group, are given in the diagnosis of the melanopus group and in the key (see below). In many characters of the structure of abdomen and male genitalia, the canaliculatus group also resembles the aggregate including the large and diverse atratulus -fuscipes -cautus -crassus complex and some isolated American members, as S. (N.) mendosus Puthz, 1971 ( 1971d) and S. (N.) sordidus Puthz, 1988, but phylogenetic relations within the named aggregate remain poorly understood; therefore a thorough analysis seems to be premature. In external morphology, the canaliculatus group can easily be distinguished from all the species of the mentioned complex by the presence of the long and sharp, line-shaped, longitudinal median furrow of the pronotum.
Taxa included
S. (N.) alopex sp. nov., S. (N.) brivioi Puthz, 1972 , S. (N.) canaliculatus Gyllenhal, S. (N.) canalis sp. nov., 1827, S. (N.) canosus sp. nov., S. (N.) caseyi Puthz, 1972 , S. (N.) confusus J. Sahlberg, 1876 , S. (N.) delitor sp. nov., S. (N.) dolosus Casey, 1884 , S. (N.) geminorum Puthz, 1973 * (USA: Washington; Canada: BC, see Puthz 1973b: 208), S. (N.) idoneus Sharp, 1887 * (Mexico, see Sharp 1887: 799; Puthz 1968: 11 <as laniger Puthz, 1968 >), S. (N.) illotulus Puthz, 1972 , S. (N.) illusor Ryvkin, 1987 , S. (N.) immigratus Puthz, 2006 * (China: Taiwan: Ilan Hsien: Shen Mi Lake, 1100 m, see Puthz, 2006: 187), S. (N.) labilis Erichson, 1840 , S. (N.) latipennis J. Sahlberg, 1880 , S. (N.) milleporus Casey, 1884 , S. (N.) nitens Stephens, 1833 , S. (N.) raddei Ryvkin, 1987 , S. (N.) shogun Puthz, 1987 * (Japan: Nishigo Uzan, see Puthz 1987: 45), S. (N.) sphaerops Casey, 1884 , S. (N.) vinnulus Casey, 1884 .
Notes on comparative morphology and evolution
As most other polytypic groups in Steninae , the canaliculatus group reveals diverse evolutionary trends quite evident at a comparison between species of the same lineage. A concise review of the mentioned trends is given below.
A. Upper surface of head between eyes tends either to become flattened, at most slightly convex ( S. shogun , S. nitens , S. illotulus , S. raddei , S. caseyi , S. canaliculatus , S. canalis , S. canosus , S. alopex ) or to produce distinct longitudinal impressions with median elevation in between ( S. labilis , S. idoneus , S. vinnulus , S. brivioi , S. confusus , S. delitor , S. illusor , S. latipennis , S. immigratus , S. geminorum , S. sphaerops , S. dolosus , S. milleporus ). The latter tendency seems to arise and develop independently, at least within the complexes labilis-confusus-delitor-illusor-latipennis-sphaerops and dolosus - milleporus , in spite of the fact that the latter is closely related in the shape of the male genitalia to the ‘ canaliculatus s.str. ’ complex characterised by a flat front.
B. For three species studied by him ( S. canaliculatus , S. nitens , S. labilis ), Betz (1996) has shown a considerable reduction of the adhesion capture apparatus. It is expressed both in the degeneration of sticky cushions of paraglossae (decreasing of their surface and number of adhesive setae etc.) and in a shortening of the labium. No data on labium length have been provided by the named author but, as it follows from the text, S. milleporus (as sectilifer Casey, 1884 ), S. caseyi , S. vinnulus , as well as S. melanopus , which is regarded as a member of the same group, have also been studied in this respect.
I have measured the relative length of the prementum (eulabium after Weinreich 1968) for six species of the canaliculatus group: S. canaliculatus (I: 17, 0.53; II: 16, 0.53), S. nitens (33, 0.87), S. labilis (37, 1.12), S. illotulus (26, 0.84), S. raddei (35, 1.09), S. latipennis (31, 0.79); and for three species of other groups of the subgenus Nestus Rey, 1884 : S. ruralis Erichson, 1840 (47, 1.52) from the palposus group, S. argus Gravenhorst, 1806 (47, 1.74) from the fuscipes group, and S. melanopus Marsham, 1802 (41, 1.46) presumably representing a monotypic group. In each case above, the first number in brackets means length of prementum whereas the second one is a rounded ratio between the former and the length of the head (two specimens have been measured for S. canaliculatus ). One can see that the data split the examined species into three clusters: a) S. canaliculatus ; b) the remaining species of the canaliculatus group; c) the species of other groups; both total and relative length of prementum increases from category a) to c). It can be supposed that the reduction of the eulabium is a characteristic feature of all the members of the canaliculatus group, but it must be verified by a specific study. The shortening of the adhesion capture apparatus seems to correlate with some other modifications of the mouth parts, namely the more proximal position of the internal tooth of mandibles as well as its lesser deflection off the main plane of the mandible.
The small median notch of the labrum is a specific character of S. canaliculatus and the most closely related Palaearctic species ( S. canalis , S. canosus , S. alopex ). Though the functional explanation of this feature is obscure as yet, one can suppose it to correlate with the extreme reduction of the adhesion capture apparatus in this species complex. The Nearctic members of the same complex ( S. milleporus and S. dolosus ) have only a broad and shallow emargination on the anterior margin of the labrum; the margin is slightly sinuate at the middle in S. nitens , S. illotulus , S. raddei , S. caseyi , S. brivioi , S. vinnulus ; lastly, the species of the labilis-confusus-delitor-illusor-latipennis-sphaerops line have the margin nearly straight.
I believe that the modification of the mouth parts should be connected more likely to a particular suite of potential prey species. Unfortunately, Betz used only springtails and (to a lesser extent) winged aphids in his interesting prey-capture experiments, but those may not exhaust the list of possible prey objects for Steninae . I did not experiment with species of the canaliculatus group; however other Stenus species were capturing nymphs of Miridae ( Insecta: Hemiptera : Heteroptera ) and Cicadellidae ( Insecta: Hemiptera : Auchenorrhyncha ), juvenile spiders of the family Linyphiidae ( Arachnida: Aranei), small mites ( Arachnida: Acari), small pyralid caterpillars ( Insecta: Lepidoptera : Pyralidae ), small flies (Diptera), soil Enchytraeidae (Oligochaeta) , in addition to the aforementioned objects, in my unpublished experiments of 1985–1986. I consider it necessary to undertake a more detailed study of prey for the species of the group being discussed.
C. The tendency of legs to become longer has most likely taken place independently in two species complexes: delitor-illusor-latipennis-confusus-sphaerops and nitens-illotulus-raddei (only the species with the 1 st segment of the metatarsus longer than the 5 th are listed). S. canaliculatus , S. canalis , S. canosus , S. alopex , S. milleporus , S. caseyi have metatarsi with the 1 st segment about as long as 5 th; in S. dolosus , S. brivioi , S. vinnulus , and S. labilis , metatarsi remain short (with the 1 st segment evidently shorter than the 5 th).
D. The plesiomorphic aedeagus with a lanceolate median lobe and unmodified cylindrical parameres (as in S. labilis ) can be transformed by different ways. The distal sclerotized part of the median lobe may be shortened, either angularly (as in S. sphaerops ) or rounded-angularly (in the delitor-illusor-latipennis line); in only one case with a small apical denticle (in S. vinnulus ); this modification usually correlates to a lengthening of the whole aedeagus (see Figs. 6 View Fig , 7 View Fig A–I) or only the median lobe (as in S. confusus , see Fig. 2E View Fig ), although this correlation has not been observed in S. raddei , belonging to another lineage ( Fig. 2A, B View Fig ). The tendency of the median lobe to become narrower (as in S. shogun , S. brivioi , S. alopex ) is another way of transformation, being peculiar to the lineages of nitens and ‘ canaliculatus s.str. ’; one can suppose that the aedeagus of Mexican S. idoneus has evolved in this way. All the modifications mentioned may be followed by forming a more or less developed medial impression or/and keel at the distal sclerotized part of the median lobe.
The main trend in the evolution of the parameres is a differentiation of their setiferous surface. It is expressed more frequently in a flattening of the apicointernal side of each paramere or, on the contrary, in forming a one-sided swell which setae are attached to.
Based on all the characters and tendencies briefly discussed above, we can subdivide the canaliculatus group into four subgroups: the monotypic labilis subgroup ( S. labilis ), the nitens subgroup ( S. immigratus , S. nitens , S. raddei , S. illotulus , S. shogun , S. caseyi , S. geminorum , S. brivioi , S. idoneus ), the canaliculatus s.str. subgroup ( S. dolosus , S. milleporus , S. canaliculatus , S. canalis , S. canosus , S. alopex ), and the confusus subgroup ( S. confusus , S. delitor , S. illusor , S. latipennis , S. sphaerops , S. vinnulus ).
Bionomics
The majority of the species for which enough data have been presented can be grouped into swamp inhabitants and alluvial ones. The former category, comprising the species dwelling in moss and litter of eutrophic and mesotrophic wet habitats, definitely includes S. nitens , S. raddei , S. illotulus , S. caseyi , S. brivioi and S. sphaerops (?), whereas the latter category consists of the members inhabiting open alluvia, like shingle beds, sandy, clayey or silty banks, sometimes sparsely overgrown with grass but without any more or less evident peat-moss or litter layer: S. labilis , S. delitor , S. latipennis , S. confusus , S. dolosus , S. canaliculatus , S. canosus , S. canalis . There are also some species ( S. alopex , S. vinnulus , and supposedly S. illusor ) living in biotopes of both categories. The peculiar swamp species, excluding S. sphaerops for which too little data are available, are represented only by the nitens subgroup, which is supposed to be closest to the common ancestor of the canaliculatus group.
General distribution
Palaearctic except subtropical regions (spotty-relict in the southernmost temperate territories); the whole Nearctic; the Caribbean Islands. Many species of the group are represented in the northernmost territories of the Holarctic.Thus, five of the fifteen Palaearctic species have been recorded for the Taimyr Peninsula and Putorana Highland, which is more than 1/4 of the regional stenine fauna.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Steninae |
|
Genus |
|
|
SubGenus |
Nestus |