
Chaltenobatrachus grandisonae ( Lynch 1975 )
|
publication ID |
https://doi.org/10.5281/zenodo.202477 |
|
DOI |
https://doi.org/10.5281/zenodo.5620243 |
|
persistent identifier |
https://treatment.plazi.org/id/823487A8-A10F-FF9D-FF50-F937FBCDFEF8 |
|
treatment provided by |
Plazi |
|
scientific name |
Chaltenobatrachus grandisonae ( Lynch 1975 ) |
| status |
comb. nov. |
Chaltenobatrachus grandisonae ( Lynch 1975) comb. nov.
Telmatobius grandisonae Lynch (1975) Atelognathus grandisonae Lynch (1982) Alsodes monticola Cei and Gil (1996)
New specimens. CNP A–392, 393, 342, 343, 394–400 collected 27 January 1997, Lago del Desierto, Santa Cruz Province, Argentina ( 49º04’41’’S, 72º54’17’’W).
Description (based on types and new available specimens). Small to medium frog (adults up to 46 mm SVL; Table 1 View TABLE 1 ) with frog–like appearance, body and limbs of regular proportions. Head somewhat smaller than one third of body length, depressed, rounded in dorsal view, wider than long. Snout shorter than ocular diameter, rounded to slightly truncate in dorsal view; sloping, with a noticeable angle at the level of the nostrils in lateral profile. Upper lip slightly protruding over lower lip. Nostrils oval, somewhat protuberant, directed dorsolaterally, approximately equidistant between the tip of the snout and the eye. Canthus rostralis rounded, loreal region weakly concave and strongly sloping towards the lip. Eyes large, very prominent and fairly lateral (in dorsal view protruding from the mandibular branches), oriented laterally. Pupil rhomboidal; a small and round ventral pupillary nodule on lower border of iris evident in some specimens. Interocular distance smaller than upper eyelid width, and similar to internarial distance. Tympanum absent. Supratympanic fold noticeable and thick, extending from the posterior end of the eye to the insertion of the arm. Tongue rounded, somewhat emarginate posteriorly (notched in exemplar MLP A–5259), free laterally and posteriorly. Premaxillary and maxillary teeth large; vomerine teeth in two oval, prominent patches, located close together, between and at posterior level of the choana. Choanae rounded, not concealed by the pars palatine of maxilla. Arms thin and of moderate length. Fingers moderately long, with noticeable interdigital membrane, more developed between the pollex and second finger and between the second and third fingers; tips of fingers rounded ( Figure 3 View FIGURE 3 a). Pollex very wide at its base. Length of fingers in increasing order: 1=2–4–3. Metacarpal tubercles large but low, the inner oval, the external weakly bifid; subarticular tubercles large and rounded or wider than long, the proximal ones larger and more protuberant; supernumerary palmar tubercles absent. Forearms somewhat more robust than arms. Legs thin and of moderate length. Toes depressed, with thick interdigital membrane, deeply incised, continuing as a dermal fringe on all toes until almost the rounded tip. Toe V with narrow dermal fringe. Length of toes in increasing order: 1–2–5–3–4. Plantar skin turgid; two little, low, noticeable metatarsal tubercles ( Figure 3 View FIGURE 3 a), the inner one oval and the outer one smaller and circular; subarticular tubercles small, rounded; supernumerary plantar tubercles absent. Tarsal fold noticeable, extending along most of the tarsus and continuous with the dermal fringe of the first toe. Tibiotarsal articulation reaching the eye. When femurs and tibias placed at right angle to the sagittal plane, the tibiotarsal articulations not in contact. Skin thin and mucoid. Back of the head, dorsum, and legs finely granulated with numerous warts, soft and turgid, of variable size and shape (rounded or elongated). Ventral skin smooth and loose; seat patch with turgid vascularized warts, restricted to the lower side of the thighs near the vent. Gular fold faint. Males with nuptial excrescences in the form of asperities, little pigmented, wide on the dorsal and inner side of the pollex and less extensive on the inner side of the second finger; also reaching the inner edge of the inner metacarpal tubercle.
Color in life. Back of the body and limbs fairly uniform bright green color, with brown to reddish warts (Figure 2). Dark, diffuse transverse bands on the dorsal side of the legs, more evident in juveniles. Belly and lower part of the flanks greyish, evenly speckled with tiny black spots. Throat whitish speckled with small dark spots. Ventral skin of the legs brownish and translucent. A diffuse dark brown band from the tip of the snout to the eye, including the nostrils, continuing to the axillary region along the supratympanic fold. A dark patch under the eye reaching the upper lip. Iris orange with gold spots; a dark vertical line under the pupil. Metamorphs show the typical species coloration. Juveniles with more intense colouring of the transverse bands of the legs, the dorsal warts and the bands on the head.
Osteology. (Description based on CNP A–392, adult male). Skull as long as wide ( Figure 3 View FIGURE 3 b). Frontoparietals paired, moderately extensive, exposing a thin frontoparietal fontanelle (frontoparietals nearly contacting each other in holotype), widely separated from nasals. Nasals relatively small, separated from one another medially and from the pars facialis of the maxilla laterally. Maxillary arch incomplete, lacking independent quadratojugal. Alary processes of premaxillae extensive, directed dorsally. Pars dentalis of premaxilla bearing 9−11 well developed pedicellate teeth; pars palatina narrow, with prominent palatine process. Pars facialis of maxilla rectangular; pars dentalis bearing 26−28 teeth extending up to the middle of the orbit; pars palatina narrow, no pterygoid processes developed. Parasphenoid triradiate; cultriform process lanceolate, not reaching anteriorly the level of the planum antorbitale; parasphenoid alae directed laterally. Vomers relatively small, widely separated medially; dentigerous processes situated between and partially posterior to choanae, each bearing 2−3 well developed teeth; prechoanal, postchoanal, and anterior processes of vomer well developed; anterior processes widely separated from the maxillary arcade. Palatines relatively long, reaching the maxilla, bearing well developed anterior processes. Pterygoids triradiate; anterior ramus of pterygoid in contact with maxilla, not reaching the planum antorbitale; medial ramus of pterygoid in contact with otic capsule but not bearing a bony articulation. Zygomatic rami of squamosals relatively long; otic rami shorter than zygomatic rami, overlapping the cartilaginous cristae parotica. Sphenethmoid well ossified, extending anteriorly notably beyond the anterior edge of nasals, with an anteromedial ossification between the nasal capsules; bony sphenethmoid not reaching the anterior margin of optic foramen posteriorly. Prootics fused with exoccipitals; epiotic eminences well developed. Occipital condyles lacking a constricted base and slightly separated medially. Opercula present, not mineralized. Columellae and tympanic annuli absent.
Hyoid plate width approximately equal to medial length ( Figure 3 View FIGURE 3 c); hyalia long, lacking anterior processes; anterolateral and posterolateral hyoid processes present, long and slightly expanded distally; posteromedial hyoid processes long and well ossified.
Eight procoelous, nonimbricate presacral vertebrae. Cotylar facets of atlas narrowly separated, approaching a Type II condition ( Lynch 1971). Transverse processes of presacrals II −IV broader than those of vertebrae V −VIII. Transverse processes of presacrals II, VII and VIII distinctly oriented anteriorly; processes of presacrals V–VIII slightly shortened. Sacral diapophyses slightly deflected posteriorly; moderately dilated. Sacrococcygeal articulation bicondylar. Urostyle with small dorsal crest on proximal half.
Pectoral girdle arciferal ( Figure 3 View FIGURE 3 d). Omosternum cartilaginous; elongate, with anterior end expanded. Scapula bicapitate. Clavicle arched, not overlaying the pars acromialis of the scapula. Coracoid narrowly dilated at its distal and proximal ends. Sternum cartilaginous, expanded posterolaterally. All cartilage of pectoral girdle calcified to some degree in adult specimen examined. Cleithrum cleft distally.
Ilial shaft cylindrical, elongate; dorsal crest poorly developed; dorsal prominence directed dorsolaterally; protuberance evident; preacetabular angle approximately 90°. Ischia small, lacking prominent processes. Pubis cartilaginous, calcified.
Humerus slender, bearing distinct crests; crista ventralis moderate; crista medialis slightly larger than crista lateralis. Phalangeal formula of hand 2–2–3–3. Terminal phalanges knobbed. Prepollex with two ossified elements. Inner metacarpal with a notorious distal flange at medial margin of the bone. Prehallux with one basal element ossified and three distal cartilaginous elements. Phalangeal formula of foot 2–2–3–4–3.
Tadpole (based on 6 specimens at stages 31 to 39 sensu Gosner (1960), CNP A–395–400) Medium–sized larva (up to 56 mm) ( Figure 4 View FIGURE 4 a,b; Table 2 View TABLE 2 ). Body elliptic in dorsal view, with a slight constriction posterior to the head; body 2 times longer than high, somewhat depressed (1.2 times wider than high), 35–40% of total length. Snout rounded in dorsal and lateral view. Nostrils small and circular, with an elevated internal rim, in dorsolateral position, slightly closer to the snout than to the eyes, opening into a depression, not raised. Distance between nostrils smaller than distance between eyes (0.4 to 0.6 times). Eyes large, in lateral position, oriented dorsolaterally, small umbraculum (fleshy projection of the iris over part of the pupil) present. Spiracular tube sinistral and conspicuous; the distal end free from the body wall. Spiracle posterolaterally directed, located beneath the mid-line of the body, 46–62% of body length, inner wall free. Vent tube dextral, short, with dextral opening.
Tail slightly higher than the body, about 60−65% of total length. Caudal musculature robust, not reaching the tip of the tail. Tail axis straight. Dorsal and ventral fins well developed, of similar height, with gently arched margins; dorsal fin arising from the last part of the body or at the tail-body junction; ventral fin arising at the level of the cloacal opening. Tip of the caudal fin rounded.
Oral disc subterminal, ventral ( Figure 4 View FIGURE 4 c), emarginated (with lateral constrictions). Single row of marginal papillae, with very large rostral gap (62−78% of the oral disc width). In dorsal view anterior and lateral papillae protruding from the contour of the snout. Supra-angular, angular and infra-angular intramarginal lateral papillae present, the latter conspicuous and numerous. All papillae sharp-pointed. Intramarginal mental papillae absent. Labial teeth well developed and keratinized. Labial tooth row-formula 2(2)/3(1). P1 gap very narrow. Upper and lower jaw sheaths well developed, wider than high, with finely serrate margins; lower jaw sheath strongly curved. Individuals raised through metamorphosis in the laboratory had the morphological and coloring features that allowed them to be identified as the same species. Metamorphs (stage 46) measure between 14.9 and 16.3 mm.
Color of tadpole. In life, dorsal color dark brown with golden spots ( Figure 4 View FIGURE 4 a). Ventral and lateral skin transparent, forming a distinct translucent contour around the body. Ventral surface of the head unpigmented, with chondrocranium, gills and heart visible through transparent skin. Abdomen black with few groups of golden, superficial guanophores. Caudal musculature dark brown with irregular golden patches. Fins translucent with scattered melanophores along the course of blood vessels, and scattered groups of guanophores. Iris gold, with dark spots. Nostrils with a pigmented rim. Spiracular tube transparent. Keratinized buccal structures highly pigmented.
Karyology. Chaltenobatrachus grandisonae has a diploid complement of 32 chromosomes in 87 metaphases. The analysis of 16 metaphasic plates revealed a fundamental number (FN) of 60. Pairs 7, 9, 10, 11, 12, 14 and 15 are metacentric (m); pairs 1, 6 and 8 are submetacentric (sm); pairs 2, 3, 4 and 5 are subtelocentric (st) and pairs 13 and 16 are telocentric (t) ( Figure 5 View FIGURE 5 a; Table 3). Pair 7 has a secondary constriction (SC) on the large arm in interstitial position in all plates studied. This SC shows an achromatic gap variable in size in the homologous pair. The specimens showed active ribosomal cistrons on the secondary constriction ( Figure 5 View FIGURE 5 b). When chromosomes are arranged according to their decreasing length, pairs 1 and 2 are large (>100 units), pair 3 is intermediate (between 80 and 100 units) and pairs 4 to 16 are small (<80 units) (Table 3). Thirty six percent of the length of the karyotype is taken up by the three first large-intermediate pairs of chromosomes, while sixty four percent is taken up by the thirteen small chromosomes.
head width; ED: eye diameter; END: eye-naris distance; UEL: upper eyelid length; UEW: upper eyelid width; IOD: interorbital
distance; IND: internarial distance; SED: snout-eye distance; TL: tibia length; FoL: foot length; FL: femur length. * Cleared
and stained.
Measurements Stage
Phylogenetic relationships. The DNA dataset included a total of 3018 base pairs (933bp from 12S, 1036 from 16S, 707 from cytb, and 342 from rhod), of which 384 sites were informative under parsimony. The Maximum Parsimony analysis by TNT resulted in a single most parsimonious tree of 1134 steps (ci= 0.786, ri= 0.688). In our cladogram Chaltenobatrachus grandisonae is the sister group of the genus Atelognathus . Hylorina sylvatica is sister to Batrachyla , and this clade forms a strongly supported group with the clade formed by Chaltenobatrachus + Atelognathus ( Figure 6 View FIGURE 6 ). All nodes are strongly supported. The genus Atelognathus shows a MP Jackknife value of 100 %, corroborating its monophyletic nature. For Bayesian inference analyses, the best model for our data was GTR + G. The Bayesian analyses recovery the same topology as was estimated using Maximun Parsimony in TNT, for the combined matrix as for each gene partition. The only exception is the cytb partition, where Chaltenobatrachus forms a polytomy with the monophyletic genus Atelognathus and the clade containing Hylorina and Batrachyla . All nodes in the combined matrix resulted in posterior probability values of 100%. We adopt the subfamily name Batrachylinae for the clade containing Batrachyla , Hylorina, Chaltenobatrachus and Atelognathus (see Figure 6).
Habitat and natural history. Chaltenobatrachus grandisonae lives in rain forests and wetlands of the southern Andes Mountains in Patagonia, at least from 48° to 49°S. Regarding altitude, it has been found from the level of Lago del Desierto ( 500 m a.s.l.) to very near the treeline ( 830 m a.s.l.). The type locality –plateau below the south peak ( 640 m a.s.l.), Puerto Edén on Isla Wellington– was described by Grandison (1961), extracted from a personal communication from Dr. Holdgate, zoologist with the Royal Society Expedition to Southern Chile.
The type locality and the new localities described here, Lago del Desierto and Lago Nansen, are located in a mountainous area showing strong evidence of glaciation, near large ice fields. Permanent snow and glaciers are present at the summits of many of the higher elevations. The climate in the area is cold and wet, with long winters and precipitation in the form of abundant snow.
Valleys and slopes up to about 1000 m elevation are covered by temperate-cold rain forests of austral beeches ( Nothofagus spp.). At the type locality there are tangled dwarf thickets of Nothofagus antarctica , alternating with open moorlands ( Grandison 1961). At the localities Lago del Desierto ( Figure 7 View FIGURE 7 ) and Lago Nansen ( Cei & Gil 1996) there are deciduous forests of Nothofagus pumilio with a sparse shrub stratum and a herbaceous stratum rich in pteridophytes and bryophytes, which can form dense mats covering fallen logs and standing tree trunks. In poorly drained areas around wetlands, Nothofagus antarctica is present. There are many bodies of water such as lakes of glacial origin, ponds and streams. On flat sites or those with poor drainage, communities are predominantly herbaceous hygrophytic, made up of grasses, rushes and sedges ( Gramineae , Juncaceae and Cyperaceae ), and peat bogs. Smaller bodies of water freeze superficially in winter.
TABLE 3. Centromeric Index, relative length (mean and standard deviation), and types of chromosomes of Chaltenobatrus grandisonae sp. nov. a Centromeric Index = short arm/total length of the chromosome. b Relative length according to Bogart (1970). c m: metacentric, sm: submetacentric, st: subtelocentric, t: telocentric. * Chromosomes with secondary constriction. Juveniles and adults of Chaltenobatrachus grandisonae were found under fallen logs near small water bodies, and adults were found submerged in pools, probably engaging in breeding activity. During three different summers (January) tadpoles were found in a variety of lentic environments located between 500 and 800 m a.s.l., either in wetlands or in open areas in the forest ( Figure 7 View FIGURE 7 a). These include small, shallow pools ( 3–10 m diameter, 10–20 cm depth pH = 6.5) and larger pools ( 20 m diameter, 1 m depth, pH = 6.5) with submerged macrophytes ( Miriophyllum) and spherical Nostoc sp. colonies. Tadpoles were also found in the forest in small, cold, shady pools ( 2 m diameter, 30 cm depth, pH = 6.2), surrounded by liverworts (Hepaticae) and Gunnera magellanica .
There are no data on the breeding behaviour of Chaltenobatrachus grandisonae . Mating call, amplexus type and egg laying are still unknown. Once, the release call of a female was heard. Larval development is aquatic and the larva is an active feeder. In summer (January) different sized tadpoles were found living together: small ( 18–20 mm, development stage 25), and large (up to 56 mm, stages 37 to 39). The fact that there were larvae from different cohorts present at the same time allows us to assume that they developed from clutches laid in different years, and the larvae have a prolongued development and overwinter in the water bodies. Metamorphosis must occur at the end of summer.
Chaltenobatrachus grandisonae was found sympatrically with an Alsodes species and Nannophryne variegata . Grandison (1961) also cited these two species ( Alsodes sp. cited as Eupsophus coppingeri ) for the area of the type locality on Wellington Island. In the area of Lago del Desierto, Chaltenobatrachus grandisonae tadpoles were found cohabitating the shallowest pools with Nannophryne variegata tadpoles.

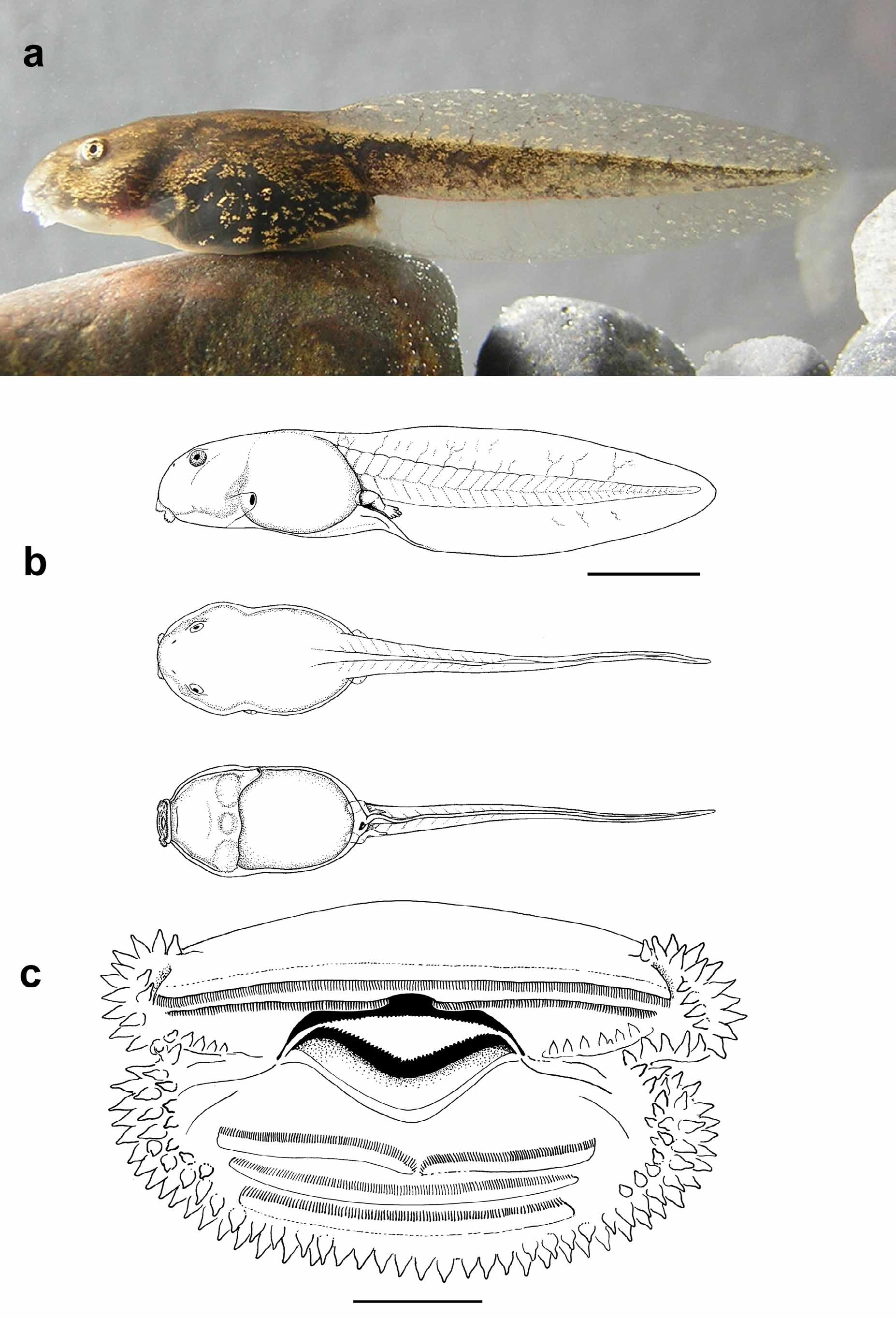



TABLE 1. Measurements of Chaltenobatrachus grandisonae adult specimens. SVL: snout-vent Length; HL: head length; HW:
| Character SVL | BM 1962.629 33.3 | CNP 209 38.2 | A– CNP 210 * 38.5 | A– CNP 211 37.5 | A– CNP 212 40.3 | A– MLP 5259 34.6 | A– MACN 36084 37.5 | Average ± SD 37.13 ± 2.39 |
|---|---|---|---|---|---|---|---|---|
| HL | 11.7 | 11.4 | 11.8 | 10.8 | 11.4 | 11.7 | 10.1 | 11.27 ± 0.62 |
| HW | 11.7 | 11.8 | 12.3 | 10.9 | 11.2 | 11.2 | 10.3 | 11.34 ± 0.66 |
| ED | 3.7 | 3.6 | 3.2 | 3.5 | 3.5 | 3.6 | 3.5 | 3.51 ± 0.16 |
| END | 2.5 | 2.7 | 2.4 | 2.5 | 2.4 | 2.9 | 2.3 | 2.53 ± 0.21 |
| UEL | 4.9 | 5.8 | 5.7 | 5.3 | 5.1 | 4.8 | 4.8 | 5.20 ± 0.42 |
| UEW | 3.6 | 2.6 | 3.3 | 2.5 | 3.2 | 3.4 | 2.4 | 3.00 ± 0.49 |
| IOD | 3.3 | 3.1 | 3.1 | 3.2 | 3.1 | 2.4 | 2.7 | 2.99 ± 0.32 |
| IND | 2.7 | 2.5 | 2.5 | 2.4 | 2.6 | 2.5 | 2.1 | 2.47 ± 0.19 |
| SED | 5.1 | 4.9 | 5.1 | 4.8 | 5.1 | 4.6 | 4.5 | 4.87 ± 0.25 |
| TL | 14.6 | 15.2 | 15.2 | 15.2 | 15.8 | 15.7 | 14.4 | 15.16 ± 0.52 |
| FoL | 21.8 | 24.3 | 23.7 | 25.1 | 24.3 | 24.4 | 22.2 | 23.69 ± 1.23 |
| FL | 15.2 | 16.8 | 17.2 | 17.4 | 14.1 | 17.4 | 14.5 | 16.09 ± 1.44 |
TABLE 2. Average ± standard deviation of the morphometric variables of Chaltenobatrachus grandisonae larvae. Stages were assigned according to Gosner (1960). Measurements are expressed in mm.
| Total length Body length | 31 (n=1) 39.5 16.2 | 37 (n=1) 49.8 19.5 | 38 (n=1) 50.2 18.9 | 39 (n=3) 49.25±1.72 17.94±0.56 |
|---|---|---|---|---|
| Tail length Maximum body height | 23.3 8.8 | 30.3 10.3 | 31.3 8.4 | 31.31±1.65 8.91±0.27 |
| Maximum tail height Caudal musculature height Maximum body width Eye diameter Interorbital distance Extraocular distance | 7.8 2.8 10.2 1.3 3.4 6.3 | 10.5 3.4 11.6 1.7 3.8 7 | 10.3 3.16 11.0 1.6 3.5 6.8 | 9.2±0.55 3.42±0.09 11.04±0.3 1.85±0.15 4±0.3 7±0.1 |
| Body width (eye level) Nostril diameter Internarial distance Extranarial distance | 8.2 0.3 2.1 2.5 | 9.9 0.2 2 2.7 | 9.51 0 3 2 2.5 | 9.57±0.26 0.26±0.04 2.05±0.13 2.4±0.41 |
| Body width (nostril level) Rostronasal distance | 5.5 1.7 | 6.5 1.7 | 5.6 1.6 | 6.9±0.66 1.65±0.24 |
| Orbitonasal distance Rostroorbital distance Rostro-spiracular distance Oral disc width | 1.5 3.7 8.3 4.2 | 2.3 4 9.0 4.4 | 1.9 4 10.5 4.6 | 2.25±0.13 4±0.47 10.55±0.66 4.5±0.05 |
| Rostral gap width Suprarostrodont length | 2.7 1.9 | 3.3 2.2 | 3.2 2 | 2.9±0.33 1.95±0.1 |
| Infrarostrodont length | 1.5 | 2 | 1.5 | 1.7±0.05 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chaltenobatrachus grandisonae ( Lynch 1975 )
| Basso, Néstor G., Úbeda, Carmen A., Bunge, María M. & Martinazzo, Liza B. 2011 |
Alsodes monticola
| Cei and Gil 1996 |
Atelognathus grandisonae
| Lynch 1982 |
Telmatobius grandisonae
| Lynch 1975 |