
Overosaurus paradasorum, Coria & Filippi & Chiappe & García & Arcucci, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3683.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:C1C6988A-6E7B-449E-9CEF-376FC9B3DCA9 |
|
persistent identifier |
https://treatment.plazi.org/id/8275F466-FFAA-6402-F496-32F84810FF2F |
|
treatment provided by |
Felipe |
|
scientific name |
Overosaurus paradasorum |
| status |
gen. et sp. nov. |
Overosaurus paradasorum gen. et sp. nov.
Holotype. MAU-Pv-CO-439 ( Figs 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 , Tables 1–4) representing an adult individual based on the closure of neurocentral sutures throughout the preserved vertebral column, the fusion of cervical ribs to their respective vertebrae, and the co-ossification of sacral vertebrae. The preserved skeleton consists of forty articulated vertebrae, including the last four cervical vertebrae, ten dorsal vertebrae, six sacral vertebrae, and twenty caudal vertebrae, the ribs of the last three cervical vertebrae, six right dorsal ribs (dorsal vertebrae 2–5, 8, 9), five left dorsal ribs (dorsal vertebrae 2–5 and 8); proximal ends of dorsal ribs of both sides of dorsal vertebrae 9 and 10, the complete right ilium and fragments of the left ilium.
Etymology. Overosaurus alludes to the Cerro Overo, the geographical provenance of the holotype; paradasorum , in recognition of Carlos Parada and his family, who have been always generous, helpful and supportive of our work in the Rincón de los Sauces area.
Geographic provenance. Cerro Overo , 40 km southwest of Rincón de los Sauces, Neuquén Province, Argentina. The site is in the vicinity of the following coordinates: S 37° 36’, W 69° 18 °; for security purposes, we refrain from providing the full GPS provenance GoogleMaps .
Geological horizon. Basal levels of the Anacleto Formation, Campanian ( Dingus et al. 2000).
Diagnosis. A small lithostrotian with the following unique combination of characters: posterior cervical vertebrae with long pre- and postzygapophyses that project beyond the anterior and posterior borders of the centrum, respectively; postspinal lamina absent in all dorsal neural spines; wide and massive 9 th and 10 th caudal centra that are slightly excavated laterally and have relatively flat ventral surfaces; laminar projection on the posterior border of the second and third dorsal ribs; ilium proportionally shorter anteroposteriorly and taller dorsoventrally than in other lithostrotians; and the preacetabular process of the ilium strongly deflected laterally and with a ventrolaterally tapering end.
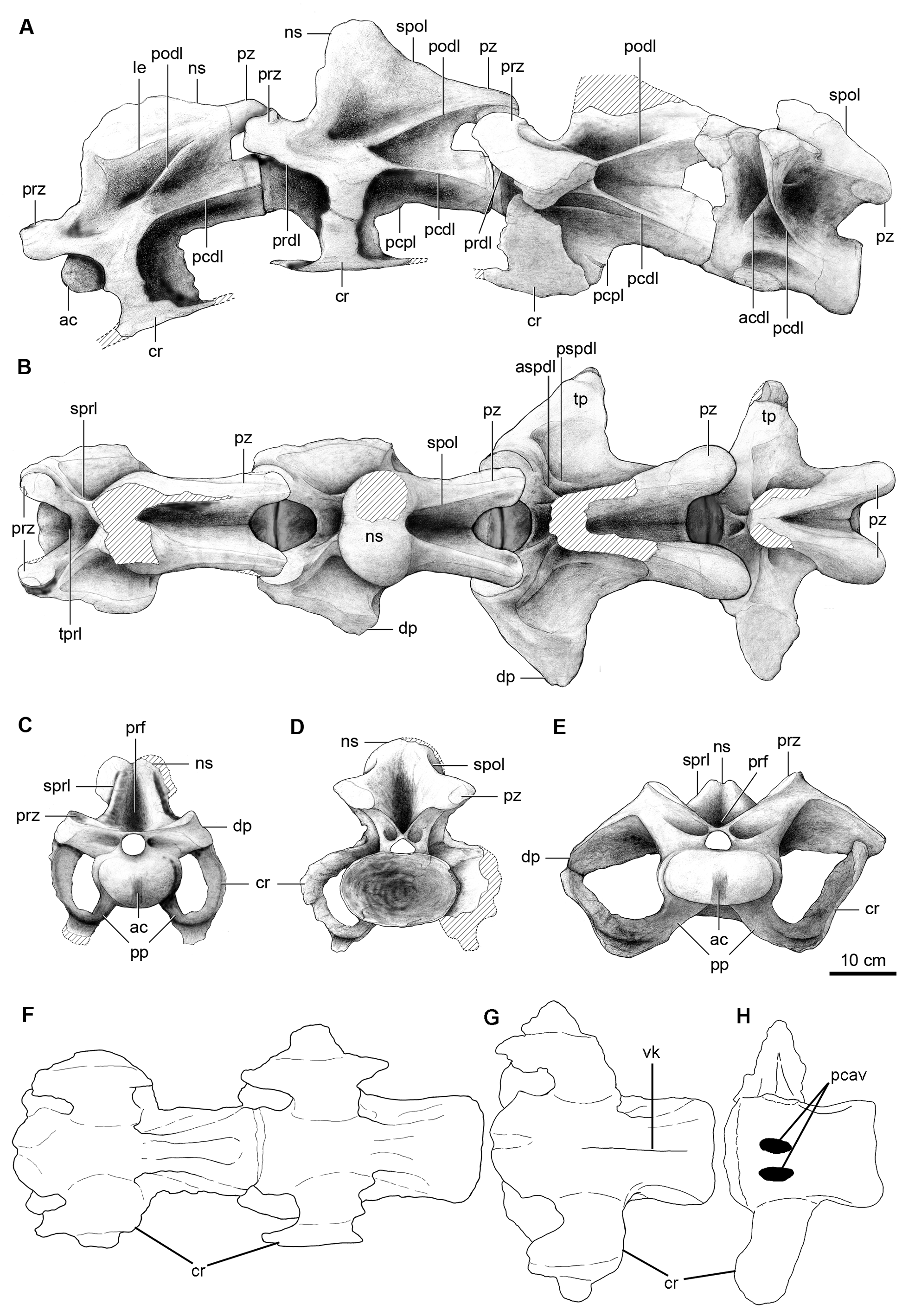
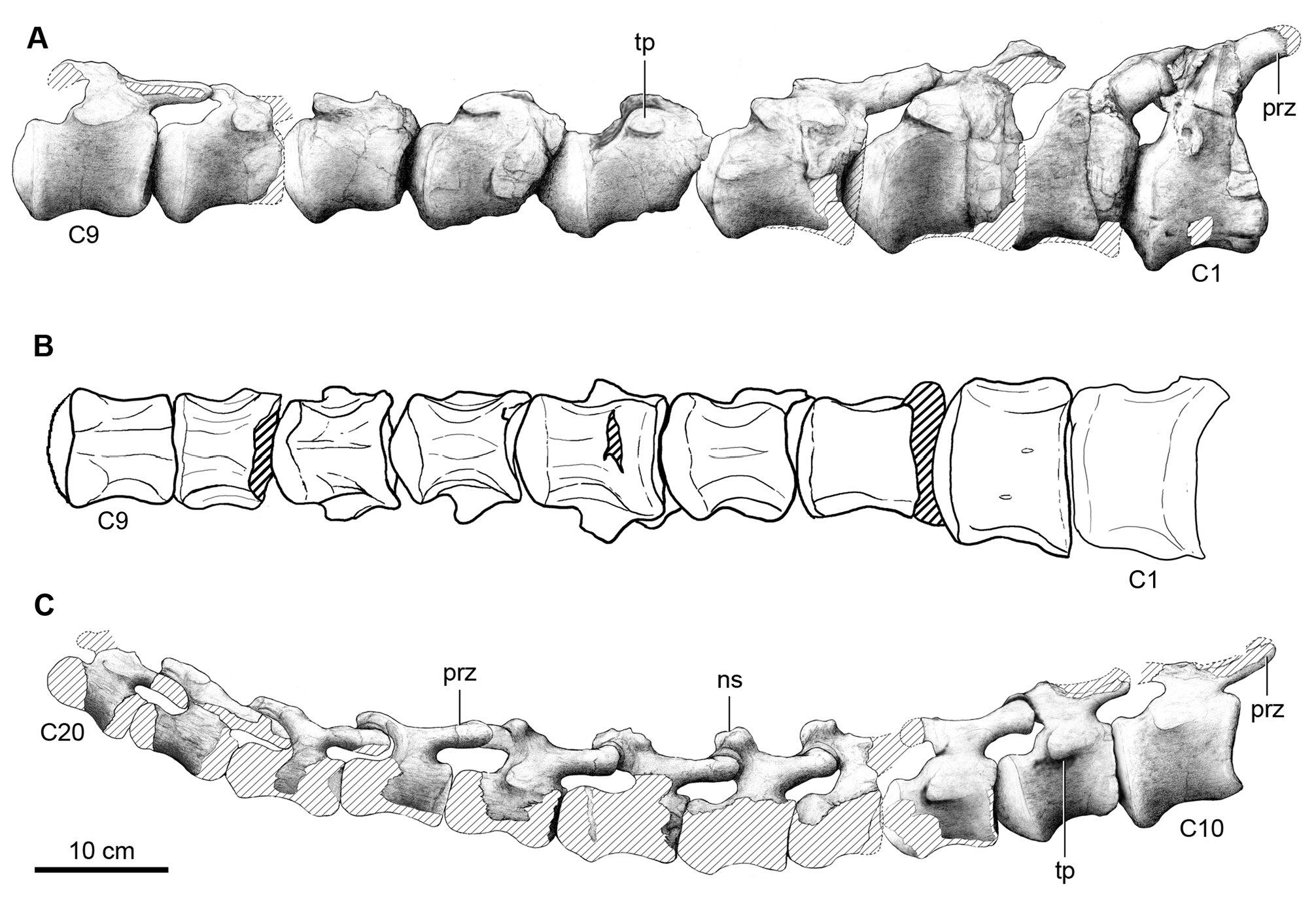
Description. Cervical vertebrae. Only the last four vertebrae of the neck, here interpreted as the 10 th, 11 th, 12 th and 13 th cervical vertebrae, are preserved ( Fig. 2 View FIGURE 2 ). Such identification is supported by comparisons with available titanosaur cervical series (e.g. MAU-PV-AC-01, Calvo et al. 1997; DGM Serie A, Powell 2003) although the vertebral numbering is tentative since the cervical series in titanosaurs could reach a total of 17 vertebrae (e.g. Rapetosaurus, Curry Rogers 2009 ). Our interpretation of these vertebrae as cervical vertebrae is based on the position of the parapophyses, located at the ventral border of the centrum and not above it as in the first dorsal vertebra, and from comparisons with other specimens with well known presacral series (e.g. Camarasaurus, McIntosh et al. 1996 ; Trigonosaurus, Campos et al. 2005 ). Weathered areas on the ventral sides of the vertebrae expose the inner somphospondylous pneumaticity.
The preserved cervical vertebrae of Overosaurus are largely complete ( Table 1), with robust and bulbous neural spines, and long, low and strongly opisthocoelous centra with deeply excavated lateral sides. The diapophyses and parapophyses are fused to the cervical ribs. The centra have an articular condyle with a conspicuous dorsoventral groove-like depression similar to that in Rinconsaurus caudamirus Calvo & González Riga (MAU-Pv-CRS02). The centra bear deep and wide lateral pleurocoels, and as in the centra of Trigonosaurus ( Campos et al. 2005) and DGM Serie A ( Powell 2003), the pleurocoels lack internal septa, which are present in Mendozasaurus neguyelap Gonzalez Riga , Saltasaurus ( Powell 1992, 2003), Rinconsaurus ( Calvo & Gonzalez Riga 2003) and most neosauropods. The parapophyses are dorsally flat, slightly convex anteroposteriorly. They are ventrolaterally projected as in some euhelopodid somphospondilians ( D’Emic 2012) and tightly fused to the cervical ribs. In lateral view ( Fig. 2A View FIGURE 2 ), the zygapophyseal peduncles are long and extend beyond the anterior and posterior edges of the centrum. This condition differs from that of other titanosaurs in which either the prezygapophyses surpass the anterior edge of the centrum [e.g. Malawisaurus dixeyi Jacobs, Winkler, Downs & Gomani 1993 ( Gomani 2005) , Rinconsaurus ( Calvo & González Riga 2003) , Trigonosaurus pricei Campos, Kellner, Bertini & Santucci 2005 , and Uberabatitan riberoi Salgado & de Souza Carvalho 2008 ] or the postzygapophyses project beyond the centrum’s posterior edge [e.g. the Brazilian specimen DGM Serie A ( Powell 2003, pl. 14), Saltasaurus loricatus Bonaparte & Powell 1980 ( Powell 2003) ], a condition interpreted as synapomorphic of saltasaurines ( Salgado et al. 1997). Furthermore, the postzygapophyseal articular facets of Overosaurus are more laterally oriented than those in Rinconsaurus , Mendozasaurus , Pitekunsaurus macayai Filippi & Garrido 2008 , Malawisaurus ( Gomani 2005) and the specimen MAU-PV-AC-01, but less than the postzygapophyseal facets of Saltasaurus and Neuquensaurus australis Powell ( Salgado et al. 2005) .
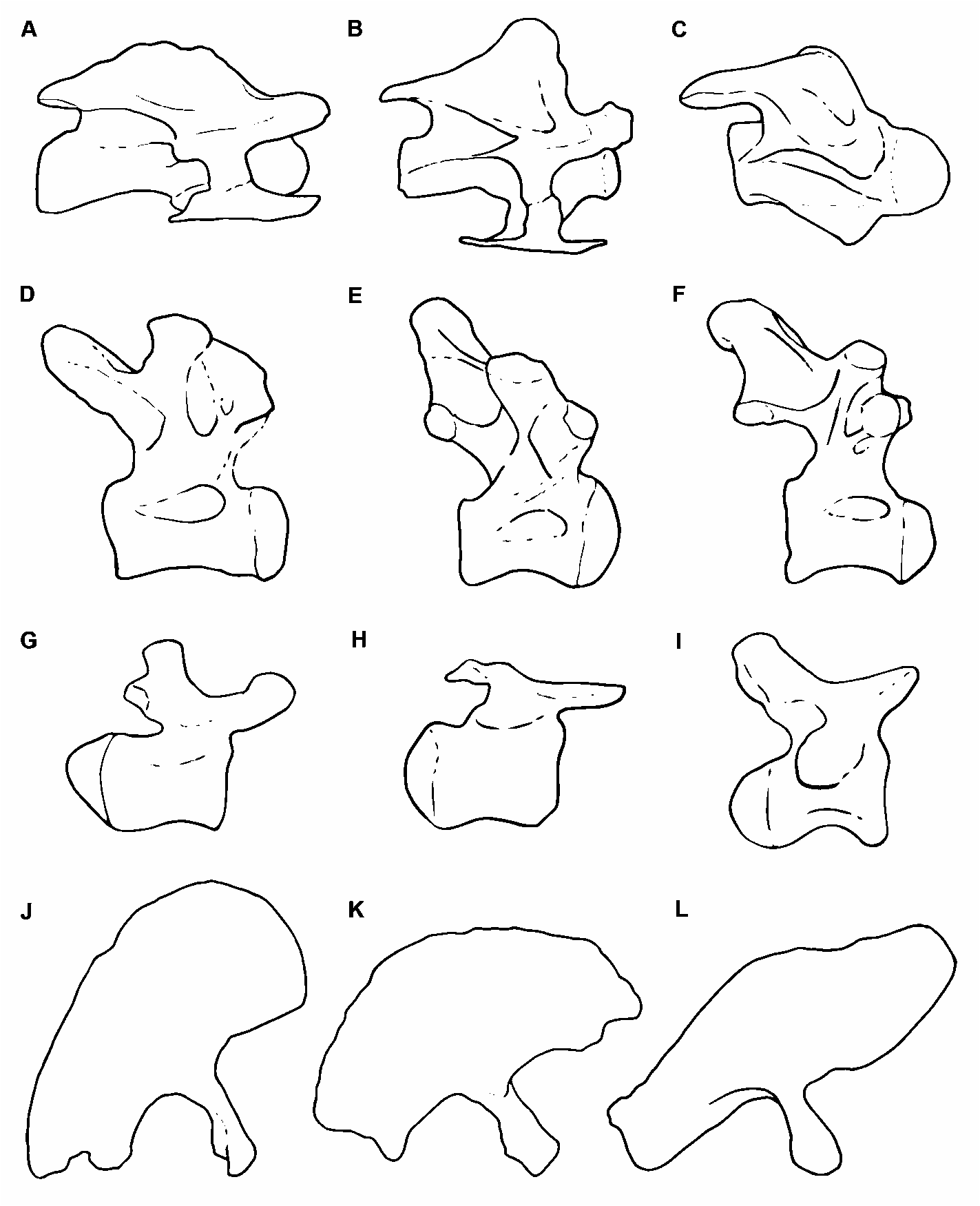
The 10 th cervical vertebra has a ventral surface of the centrum that is deeply excavated by a posterior fossa defined by two parallel ridges ( Fig. 2F View FIGURE 2 ). The neural arch has an anteroposteriorly extensive neural spine, which laterally displays a robust longitudinal edge that defines the spine’s bulbous apex. Such expansion recalls the condition of Trigonosaurus ( Campos et al. 2005) ( Fig. 8A View FIGURE 8 ) and DGM Serie A. In lateral view ( Fig. 2A View FIGURE 2 ), this longitudinal edge confines, together with the postzygodiapophyseal lamina, an extensive spinodiapophyseal fossa. As in most titanosaurs, in anterior view ( Fig. 2C View FIGURE 2 ) the spinoprezygapophyseal laminae are thick and the prezygapophyses are linked through an extensive intraprezygapophyseal lamina that forms the floor of a wide spinoprezygapophyseal fossa ( Wilson et al. 2011). The articular facets of the prezygapophyses are dorsomedially oriented. The prezygapophyses diverge laterally and their distal ends are located outside the lateral limits of the centrum, but they do not reach the level of the distal ends of the diapophyses ( Fig. 2B View FIGURE 2 ), similar to the condition in Saltasaurus ( Powell 1992, 2003) and Mendozasaurus ( Gonzalez Riga 2005) . The anterior centrodiapophyseal lamina is reduced. The transverse processes have extensive diapophyses, dorsally located above the anterior half of the centrum ( Fig. 2A View FIGURE 2 ). Above the prezygodiapophyseal laminae there is a longitudinal edge (present also in MAU- Pv-AC-01) extending over the entire diapophysis. The spinopostzygapophyseal laminae are long, well developed, and dorsally concave. Both the postzygodiapophyseal and posterior centrodiapophyseal laminae are anteriorly thin, becoming thicker posteriorly. The posterior centrodiapophyseal and posterior centroparapophyseal laminae are subparallel, reaching the posteriormost edge of the centrum. In Rinconsaurus those laminae are not parallel, but they converge posteriorly (MAU-PV-CRS03; Calvo & González Riga 2003, pl. 1), whereas in Trigonosaurus , although parallel, the posterior centrodiapophyseal lamina does not reach the centrum’s edge ( Fig. 8A View FIGURE 8 ).
The morphology of the 11 th and 12 th cervical vertebrae is similar to that of the preceding one. The ventral surface of the centrum of the 12 th cervical vertebra is concave and presents a longitudinal, medial keel ( Fig. 2G View FIGURE 2 ). These vertebrae are shorter anteroposteriorly than more anterior cervicals ( Table 1). The 11 th cervical vertebra has a more angular neural spine, in which the spinoprezygapophyseal lamina is nearly vertical ( Fig.2A View FIGURE 2 ). Furthermore, the distal bulbous end of the neural spine is less defined (the neural spine is not preserved in the 12 th cervical vertebra). The 12 th cervical vertebra has a more developed anterior centrodiapophyseal lamina, and a posterolaterally directed lamina originating at the confluence of the spinoprezygapophyseal lamina with the base of the neural spine (likely equivalent to the anterior spinodiapophyseal lamina) (see Salgado & Powell 2010).
The vertebra considered as the 13 th cervical has an intermediate morphology between the preceding vertebra and the one identified as the first dorsal. The centrum is significantly shorter anteroposteriorly than those of the preceding cervical vertebrae ( Table 1) and the diapophyses are oriented dorsally as in the dorsals. The ventral surface of the centrum has two conspicuous pneumatic cavities on either side of a broad sagittal plate ( Fig. 2H View FIGURE 2 ). The parapophyses of this vertebra are located at the very base of the centrum, as in a typical cervical vertebra, and the anterior spinodiapophyseal lamina (adjacent to the spinoprezygapophyseal lamina) described for cervical 12 is present ( Fig. 2A View FIGURE 2 ). The neural spine of cervical vertebra 13 is stouter and more strongly inclined anteriorly than in other titanosaurs (e.g. Trigonosaurus ). The prespinal fossa is small, restricted to the base of the neural spine, and not as deep as in the preceding vertebrae. As a result of the shortening of the whole vertebra, the spinoprezygapophyseal and spinopostzygapophyseal laminae are shorter and less robust.
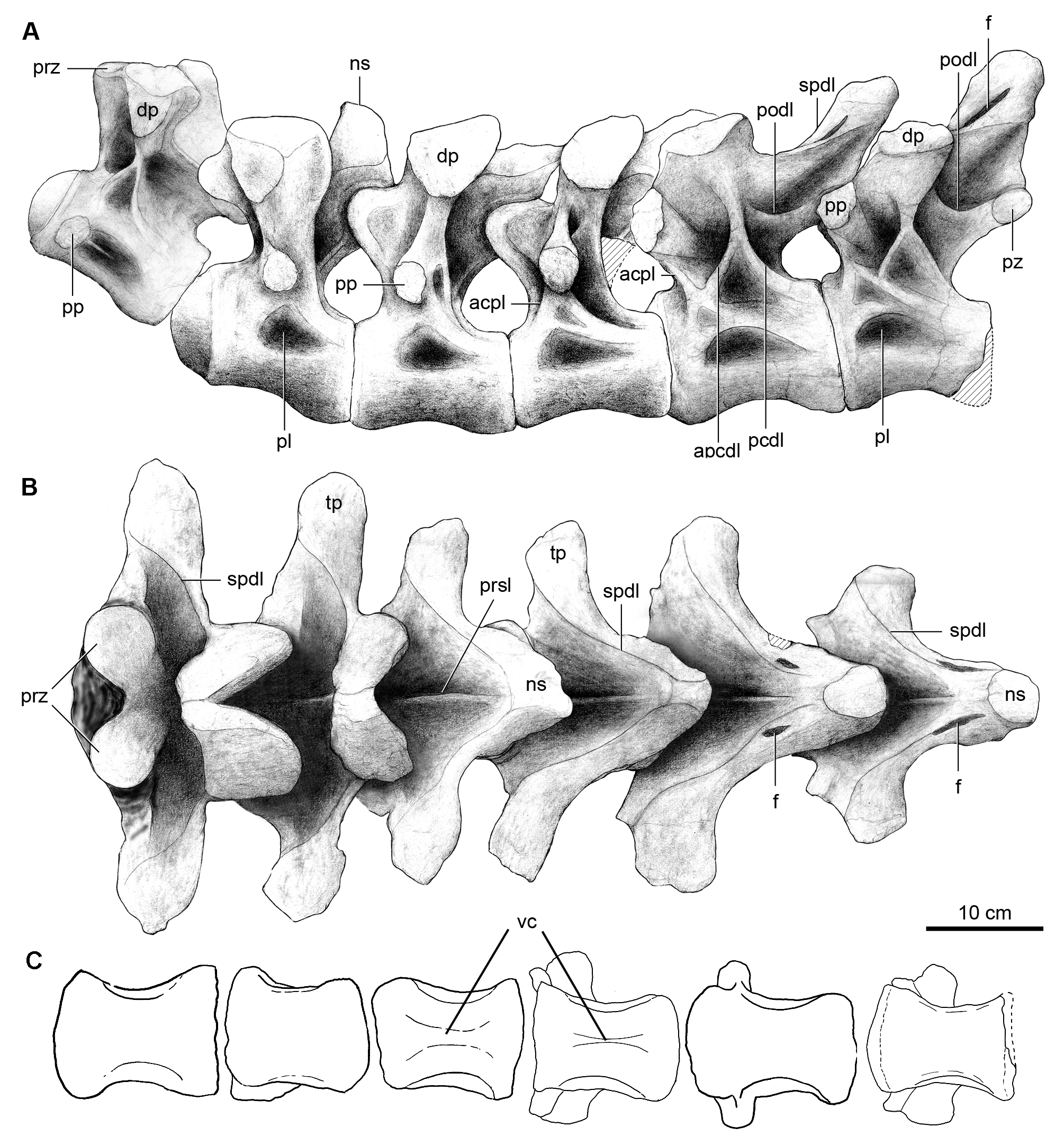
Dorsal vertebrae. The holotype specimen of Overosaurus preserves a complete dorsal series, consisting of 10 articulated and well-preserved vertebrae ( Figs 3 View FIGURE 3 , 4 View FIGURE 4 ; Table 1). Although these vertebrae are apparently not fused, they are preserved tightly articulated, which prevents the observation of their features in either anterior or posterior view. A distinctive feature of Overosaurus is the absence of postspinal laminae throughout the dorsal series. Such an absence differentiates it from Neuquensaurus ( Salgado et al. 2005) , Epachthosaurus sciuttoi Powell ( Martínez et al. 2004) , Andesaurus delgadoi Calvo & Bonaparte , Saltasaurus ( Powell 1992, 2003), Rinconsaurus ( Calvo & Gonzalez Riga 2003) , Mendozasaurus ( Gonzalez Riga 2003) , and Pitekunsaurus (MAU-Pv-AG-446), all of which possess well-developed postspinal laminae. Interestingly, the absence of postspinal laminae is shared with the basal lithostrotian Argentinosaurus huinculensis Bonaparte & Coria (MCF-PVPH-1), which in the current phylogenetic context is considered as a convergence with Overosaurus .
The centrum of the first dorsal vertebra is depressed dorsoventrally, and relatively wide mediolaterally. In ventral view, it is flat with concave sides ( Fig. 3C View FIGURE 3 ). The neural spine of the first dorsal is robust, smaller and less inclined anteriorly than that of the preceding vertebra ( Fig. 3A–B View FIGURE 3 ). It is also dorsoventrally lower than the last cervical vertebra and it possesses a deep postspinal fossa. On its anterior face, it has a prespinal lamina that extends dorsoventrally from approximately half of the height of the neural spine up to the roof of the neural canal. In lateral view ( Fig. 3A View FIGURE 3 ), the parapophysis is located immediately anterior to the lateral pleurocoel. In dorsal view ( Fig. 3B View FIGURE 3 ), the transverse processes are transversely long and somewhat narrower than those of the preceding vertebra.
The second dorsal has a mediolaterally wide and dorsoventrally depressed centrum, with its sides excavated by deep pleurocoels and a slightly concave ventral surface ( Fig. 3A,C View FIGURE 3 ). This vertebra is distinguished by having a neural spine with a broad base that narrows dorsally. The neural spine’s anterior face is relatively flat, with a gentle prespinal lamina ( Fig. 3B View FIGURE 3 ). Its posterior face bears a postspinal fossa that is shallower than that of the preceding vertebra. The lateral sides of the neural spine are tabular, anteroposteriorly broad, and vertical. The postzygapophyses are located immediately below the posterior edge of the neural spine. The parapophyses lie at the base of the neural arch.
The following two vertebrae (dorsals 3–4) have centra that are similar to the preceding vertebra, although the ventral surface of the fourth dorsal has a conspicuous longitudinal crest like Barrosasaurus ( Salgado & Coria 2009) ( Fig. 3C View FIGURE 3 ). The neural spines are thinner than those of previous vertebrae. They are strongly inclined posterodorsally ( Fig. 3B View FIGURE 3 ), as in Rinconsaurus ( Calvo & Gonzalez Riga 2003) , and have flat posterior surfaces.
The fifth dorsal vertebra has a shallow keel on the ventral surface of the centrum. The neural spine of the fifth dorsal is less inclined than in preceding dorsals. In posterior view, it has lateral expansions above the postzygapophyses. These expansions are already expressed in dorsal 4 but they acquire a greater development in dorsal 5. The apex of the neural spine of the latter is robust and roughly triangular in cross-section. Laterally on the neural spine, both anterior and posterior spinodiapophyseal laminae frame a narrow fossa. In lateral view ( Fig. 3A View FIGURE 3 ), the parapophysis has migrated to the front of the diapophysis unlike in dorsal 4 in which it was placed ventrally. The parapophysis is connected to the centrum by a short anterior centroparapophyseal lamina. The diapophysis connects with the centrum through a ventrally bifid posterior centrodiapophyseal laminae, whose posterior branch is thinner. These laminae define a triangular recess, whose ventral edge is formed by the union of the neural arch and centrum. The postzygodiapophyseal lamina is poorly developed as in Neuquensaurus and other derived titanosaurians ( Salgado 1996, Gallina 2011, Mannion & Otero 2012).
The posteriormost section of the presacral series (dorsals 6–10) is shorter, more robust, and has neural spines that are wider both transversely and anteroposteriorly. Throughout this section, the neural spines become nearly vertical and the transverse processes, short and dorsally flat, become more dorsally placed. In lateral view ( Figs 3A View FIGURE 3 , 4A View FIGURE 4 ), the bifid posterior centrodiapophyseal laminae becomes shorter towards the back. The transverse processes are shorter, dorsally flatter and more dorsally placed than in the preceding vertebrae. The postzygodiapophyseal lamina of these vertebrae is poorly developed, as in some other titanosaurs (e.g. Neuquensaurus , Trigonosaurus , Gondwanatitan ).
The ventral surfaces of dorsal centra 6 and 7 are keeled, while those of the last three dorsals are flat. The parapophyses are placed in a more anterior position and closer to the diapophysis in the seventh dorsal vertebra. In the last vertebra (dorsal 10), the parapophysis is once again ventral to the diapophysis ( Fig. 4A View FIGURE 4 ). The migration of the parapophysis results in changes to the pattern of neural arch laminae. In the seventh dorsal vertebrae the anterior centrodiapophyseal lamina is present, whereas it is absent in the ninth dorsal vertebra it is replaced by an accessory posterior centrodiapophyseal lamina. In turn, in the tenth dorsal vertebra, only the posterior centrodiapophyseal lamina is observed.
Towards the tenth dorsal vertebra, the centra are subequal in length, whereas it is shorter in the tenth dorsal vertebra.
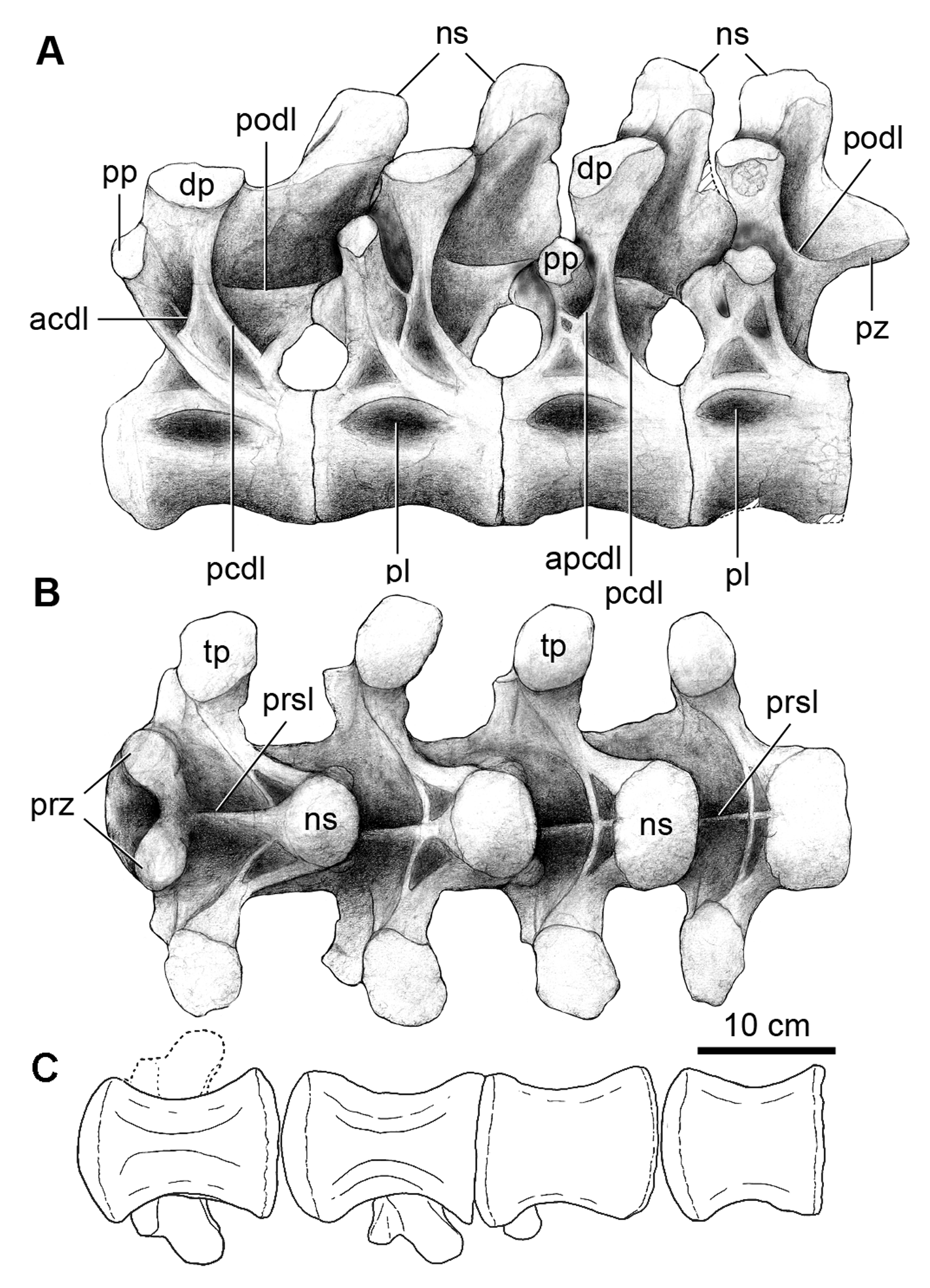
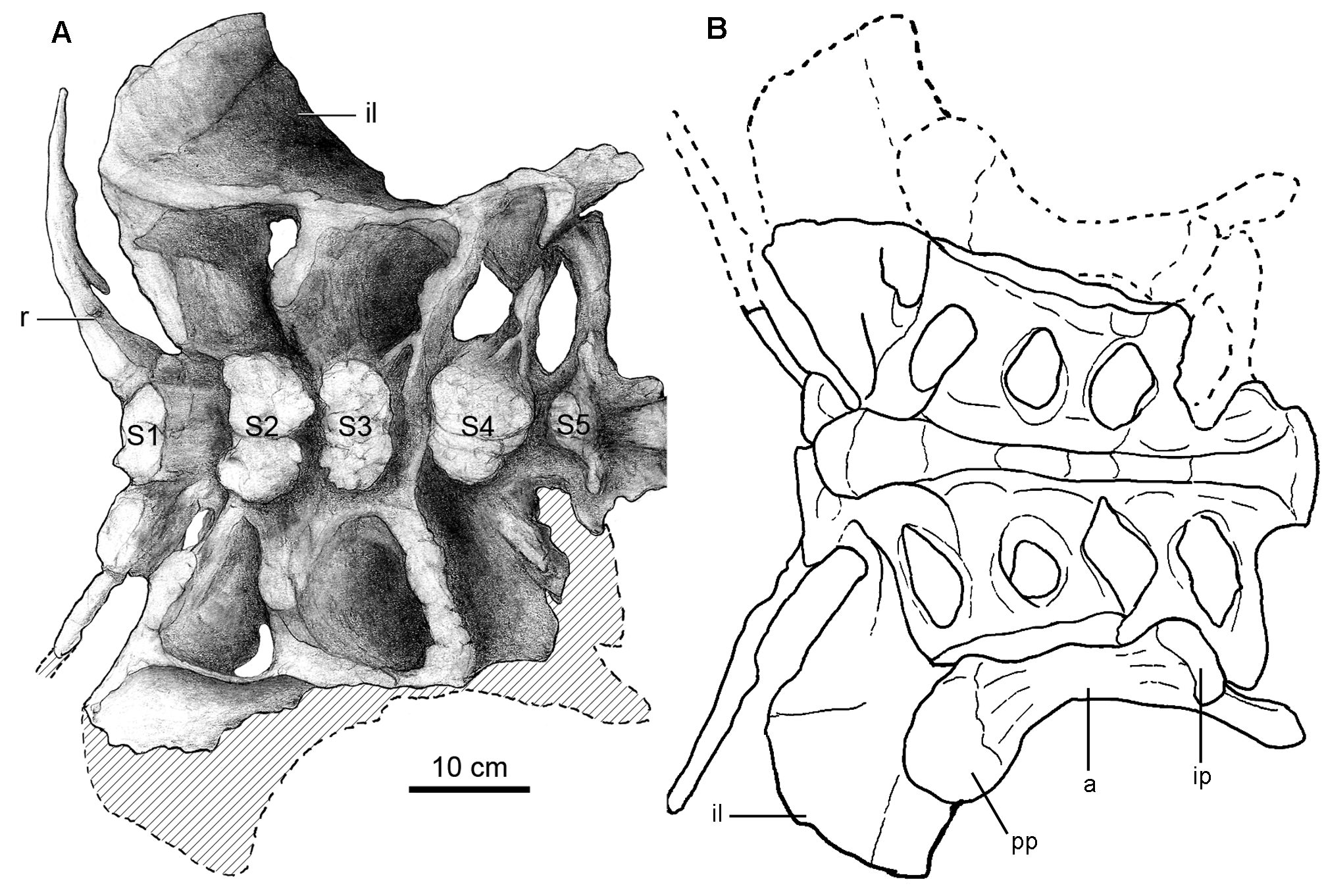
Sacrum. The sacrum is composed of six vertebrae ( Table 2) with fused centra, including a dorsosacral and a sacrocaudal vertebra as in Euhelopus ( Wilson & Upchurch 2009) , Saltasaurus ( Powell 1992, 2003), Epachthosaurus ( Martinez et al. 2004) , Argentinosaurus ( Bonaparte & Coria 1993) , and Gondwanatitan ( Kellner & Azevedo 1999) , and unlike the sacrum of Neuquensaurus ( Salgado et al. 2005) , which is composed of 7 sacral vertebrae. The dorsosacral vertebra bears a free rib, not fused to the ilium ( Fig. 5 View FIGURE 5 ). The following four sacral vertebrae have ribs and transverse processes fused to the ilium; the last sacral vertebra has its transverse processes fused to the rib of the preceding sacral and to the ilium. Ventrally, the centra are highly compressed defining a keel- like ridge ( Fig. 5B View FIGURE 5 ). The neural spines are not fused to each other but the neural arches are connected together by zygapophyseal fusion. The distal ends of the ribs of sacral vertebrae 3 and 4, and partially sacral 5, contribute to the innermost portion of the acetabulum. The sacrocaudal vertebra is more similar to the caudal vertebrae. It has longer and narrower transverse processes that abut the ilium at the level of the ischiadic peduncle. The relatively robust neural spine of this vertebra is tilted posteriorly, has a slight transverse expansion at the apex and a conspicuous postspinal lamina.
Caudal vertebrae. The preserved 20 caudal vertebrae are procoelous. While the lateral sides of caudal centra 12 –20 are somewhat weathered, the overall outline of these centra indicate that the vertebrae are procoelous throughout the available caudal section. ( Fig. 6 View FIGURE 6 ; Table 3). The caudal vertebrae of Overosaurus have centra that are transversely wide and slightly anteroposteriorly concave, with flat and broad ventral surfaces. Ventrolateral ridges are present from caudal vertebra 3. These demarcate a longitudinal ventral groove ( Fig. 6C View FIGURE 6 ). The prezygapophyses are thick. The neural spines of the first eight caudal vertebrae are missing and their neural arches are preserved up to the level of the zygapophyses. As is typical in titanosaurifoms although not exclusively, the neural arches are located above the anterior halves of the centra and they are projected in such a way that the prezygapophyses project strongly anteriorly, as in Aeolosaurus rionegrinus Powell ( Salgado & Coria 1993; Salgado et al. 1997) and Saltasaurus ( Fig. 8I View FIGURE 8 ).
paradasorum gen. et sp. nov. Measurements are in mm. Abbreviation: *, preserved.
The procoelous first caudal centrum has anteroposteriorly concave lateral and ventral sides. The ventral side is wider than the ventral side of the last sacral vertebra. The neural arch is high and anteroposteriorly narrow. The prezygapophyses are short and anterodorsally projected. The transverse processes are not preserved.
Caudal vertebrae 2–4 have procoelous and transversely wide centra, with anteroposteriorly concave lateral sides. The ventral side is wider in the second caudal vertebra, whereas it narrows on the third caudal vertebra. In turn, in the fourth caudal vertebra, the ventral side of the centrum is narrow and anteroposteriorly concave. The prezygapophyses are strongly anterodorsally projected. The transverse processes are not preserved although they seem to have been posterolaterally projected.
The 5 th –7 th caudal vertebrae have procoelous, transversely wide centra with conspicuous anteroposteriorly concave lateral sides ( Fig. 6A–B View FIGURE 6 ). The ventral sides are narrow and slightly anteroposteriorly concave. The neural spines are not preserved but the transverse processes are robust, dorsoventrally narrow, and posterolaterally projected.
The 8 th –11 th caudal vertebrae have procoelous centra, almost twice as wide mediolaterally as they are high dorsoventrally ( Fig. 6A–C View FIGURE 6 ). The lateral sides are anteroposteriorly concave, but this concavity is shallower than in the preceding vertebrae. The ventral side is almost flat and wide, mainly on caudal vertebrae 9 and 10. The prezygapophyses are anterodorsally projected. Caudal vertebra 11 preserves a dorsoventrally thick prezygapophyseal facet. A prespinal fossa is present between the prezygapophyses and the base of the neural spine. The transverse processes are reduced to a lateral swelling.
The 12 th –20 th caudal vertebrae have cylindrical and procoelous centra with anteroposteriorly concave lateral and ventral sides ( Fig. 6B–C View FIGURE 6 ). Caudal vertebrae 12–17 have rather flat lateral sides with well defined facets for the haemal arch. These vertebrae have relatively longer prezygapophyses and neural spines that are strongly inclined posteriorly, especially from the 16 th caudal vertebra. The dorsoventrally depressed condition of the saltasaurid caudal centra is not observed in the preserved caudal section of Overosaurus . On the contrary, Overosaurus exhibits the condition present in Mendozasaurus ( Gonzalez Riga 2003) and other Lithostrotia, in having caudal centra with squared, slightly concave and relatively high lateral sides (Figs. 6,8).
Cervical and dorsal ribs. The cervical ribs are strongly fused to the parapophyses and diapophyses. They have well developed cranial processes, which are triangular in dorsal view and crescentic in cross-section. The caudal processes, although preserved only on its proximal end, are long and ribbon-like ( Fig. 2A View FIGURE 2 ).
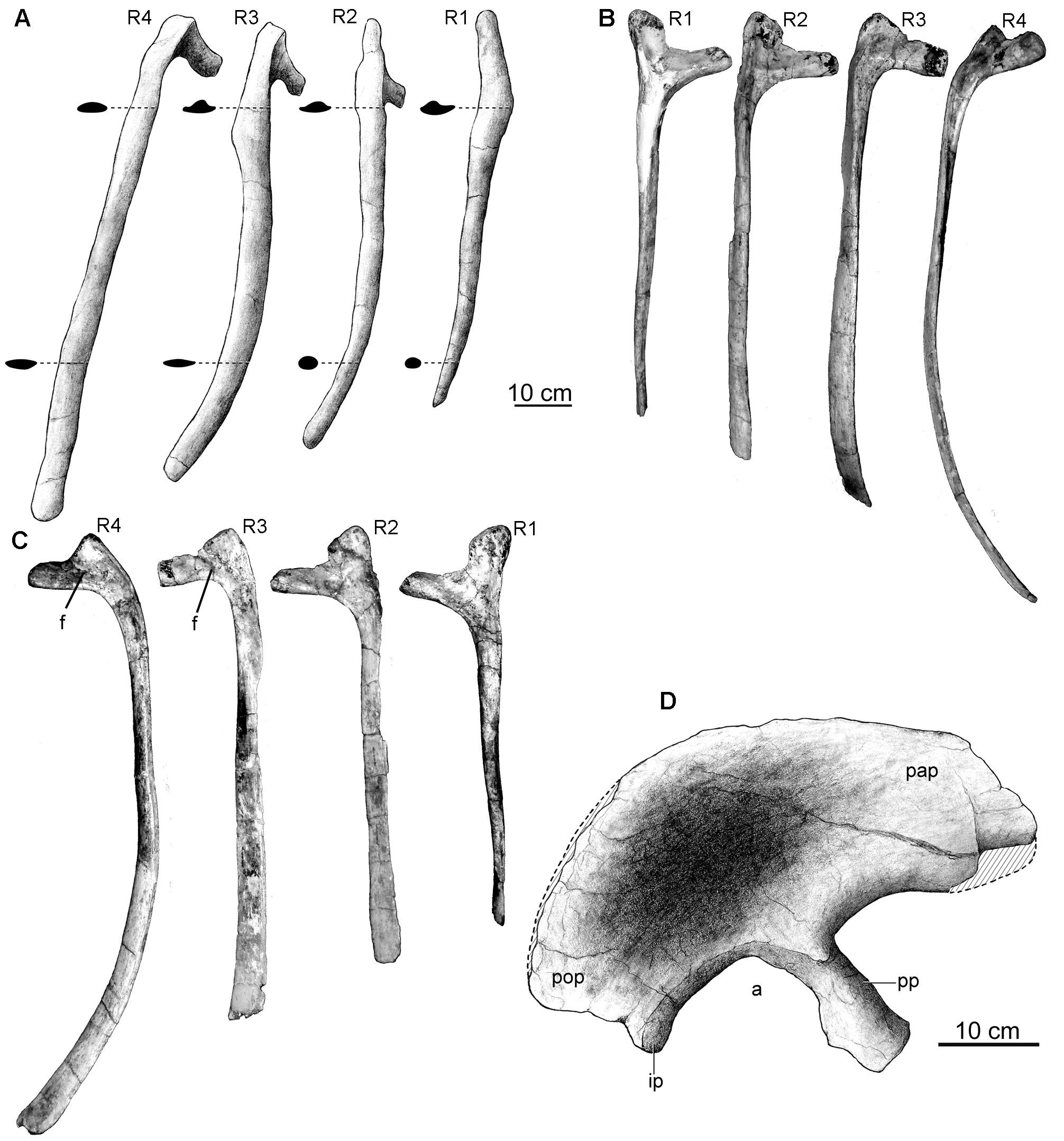
The first four pairs of preserved dorsal ribs (corresponding to dorsal vertebrae 1–4) are complete and in natural articulation ( Fig. 1B View FIGURE 1 ). They differ from subsequent ribs by being anteroposteriorly wider and more compressed, with the exception of the first pair, in which proximal and distal cross-sections are triangular and circular, respectively ( Fig. 7A View FIGURE 7 ). In lateral view, these ribs have sigmoidal outlines with distal ends curved backwards ( Fig. 7A View FIGURE 7 ). Their length increases backwards. A unique feature of the first three pairs of dorsal ribs of Overosaurus is the presence of expansions on both the anterior and posterior edges of the proximal shafts ( Fig. 7A View FIGURE 7 ). Other titanosaurs lack such expansion and their anterior dorsal ribs have parallel anterior and posterior borders.
The first pair of dorsal ribs have a capitulum twice the length of the tuberculum and are more gracile. The capitulum projects medially perpendicular to both the tuberculum and shaft. The tuberculum projects more dorsally than the capitulum. In both anterior and posterior sides, a closed depression is present between capitulum and tuberculum. The shaft is almost straight, shorter than subsequent ribs, with a triangular proximal cross-section and a subcircular distal cross-section.
The second pair of dorsal ribs has longer capitula. Both anterior and posterior depressions are present. The shaft is curved, compressed lateromedially on its proximal part and has an oval cross-section distally.
The third pair of dorsal ribs has capitula twice the length of the tubercula, which are less prominent dorsally. The posterior side bears a dorsal depression pierced by small foramina. The shaft is more curved than the second dorsal rib and it is proximal and distally laterally compressed. This rib is longer than the second one, and the expansion of the anterior and posterior borders aims its greater development.
The fourth pair of dorsal ribs has less developed capitula and tubercula. The posterior side bears a depression pierced by small foramina. It is the longest and most curved rib preserved. Unlike the tree preceding pairs, the shaft is not posteriorly curved and lacks the proximal shaft expansion.
Two complete posterior dorsal ribs (corresponding to dorsals 7–8) and parts of the last posterior ribs (dorsals 9–10) are preserved in articulation. These ribs are significantly more slender, with proximal and distal crosssections that are subcircular and elliptical, respectively. The capitulum and the tuberculum are placed at the same level in more posterior dorsal ribs, reflecting the dorsal migration of the parapophyses.
Ilium. The right ilium has an anteroposterior length of 545 mm and its height is 210 mm above the acetabulum. The iliac blade is well expanded dorsoventrally above the pubic peduncle ( Figs 7D View FIGURE 7 , 8K View FIGURE 8 ), presenting a greater dorsoventral development than in Saltasaurus and other titanosaurs ( Fig. 8L View FIGURE 8 ). The preacetabular process of Overosaurus also differs from other members of this clade in being shorter and wider, strongly deflected laterally, and having a ventrolaterally tapered end ( Fig. 7D View FIGURE 7 ). Such lateral deflection incorporates the pubic peduncle, which is located further laterally than the ischiatic peduncle. When the length of the pubic peduncle is compared to the supra-acetabular height of the ilium, the former appears shorter than in other titanosaurs. The dorsal edge of the ilium is rounded, and at the level of the acetabulum’s midpoint it descends abruptly posteroventrally ( Fig. 7D View FIGURE 7 ).
In the preserved articulation between the ilium and the sacrum, the 150 mm long pubic peduncle is perpendicular to the dorsal series. The anterior surface of the pubic peduncle is convex, the posterior one is concave, and both lateral and medial borders are roughly parallel. Transversely, the pubic peduncle is 108.7 mm wide at its proximal section, 82.5 mm wide at its mid section and 111.4 mm wide distally. The articular surface for the pubis has a kidney-shaped ventral outline and is ventromedially oriented. The acetabulum is 105 mm wide. The ischiadic peduncle is transversely wider than the surface of the acetabulum and, in lateral view, it is separated from the postacetabular process by a notch.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |