
Rhinolophus lobatus, PETERS, 1852
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zly024 |
|
publication LSID |
lsid:zoobank.org:pub:65FFC8DB-4738-49FF-99F5-FC8532CC9795 |
|
DOI |
https://doi.org/10.5281/zenodo.4328255 |
|
persistent identifier |
https://treatment.plazi.org/id/84783E54-FFA7-FFC8-AFD0-7CC1FE077115 |
|
treatment provided by |
Valdenar |
|
scientific name |
Rhinolophus lobatus |
| status |
|
RHINOLOPHUS LOBATUS PETERS, 1852 View in CoL
PETERS’ S HORSESHOE BAT
Synonyms: R. l. angolensis (Seabra, 1898), Angola, Hahna.
Type locality: Mozambique, Tete (= Tete Province), Sena GoogleMaps village, south bank of the Zambesi River GoogleMaps , - 15.679° S, 33.809° E.
Syntypes: ZMB 375 (female, poor skin only), ZMB 2496 (female, skin in alcohol, skull extracted, broken), ZMB 24922 (adult male, complete skeleton), ZMB 24927 (female, complete skeleton), collected between 1843 and 1847 by W. C. H. Peters from Mozambique, Tette (= Tete Province), Sena on the south bank of the Zambesi River, - 15.679° S, 33.809° E. GoogleMaps
Referred GoogleMaps specimens having molecular identifications: FMNH 228936 ( SMG 18959 ), male, 228937 ( SMG 18988 ), female, 228938 ( SMG 18989 ), male, 228939 ( SMG 18990 ), male, 228940 ( SMG 18991 ), female, 228941 ( SMG 18992 ), male, all collected on 2 May 2015 by S. M. Goodman, M. C. Schoeman and G. le Minter from Mozambique, Chihalatan Cave, 38.2 km Eof Inhassoro, - 21,671° S, 34,864° E. GoogleMaps FMNH: 228947 ( SMG 19022 ), female, 228954 ( SMG 19029 ), female, 228965 ( SMG 19040 ), female, all collected on 2 May 2015 by S. M. Goodman, M. C. Schoeman and G. le Minter from Mozambique, Mozambique, Inhambane Province, Malashane Cave , 39.1 km Eof Inhassoro, - 21,668° S, 34,847° E, and situated <2 km from Chihalatan Cave referred to above. GoogleMaps DM13905 (female), 13916 (female), DM 14531 (male), all collected by A. Monadjem, G. le Minter and E. Lagadec in July 2015 from Mozambique, Sofala Province, Gorongosa National Park.
Referred specimens having non - molecular identifications: DM 13894 (female), 13927 (male), 13938 (unknown sex), 13942 (unknown sex), 13943 (unknown sex), 13944 (unknown sex), 13945 (male), 14529 (male), 14531 (male), 14532 (female), collected by A. Monadjem, G. le Minter and E. Lagadec in July 2015 from Mozambique, Sofala Province, Gorongosa National Park . TMSA 49114 (female), TMSA 49117 (male), collected from Mozambique, Sofala Province, Gorongosa National Park by A. J. Guyton in July 2015 from Chitengo Camp and Cheringoma Plateau respectively. TMSA 14653 (female), 14655 (unknown sex), 14656 (female), 14657 (female), 14662 (male), 14663 (male), 14664 (female), collected 28 – 29 July 1964 from Mozambique, Tete Province, Muchena , - 15.679° S, 33.809° E. Incertae sedis: DM 8575 (male) from Mozambique, Cabo Delgado Province, Pemba Island (- 13.006° S, 40.524° E) and DM 8574 (female), from Mozambique, Sofala Province, Chinizuia Forest (- 18.977° S, 35.052° E), both collected by A. Monadjem in June 2006. TMSA 49115 (male), TMSA 49118 (male), collected from Mozambique, Sofala Province, Gorongosa National Park by A. J. Guyton in July 2015 from Chitengo Camp and Cheringoma Plateau , respectively.
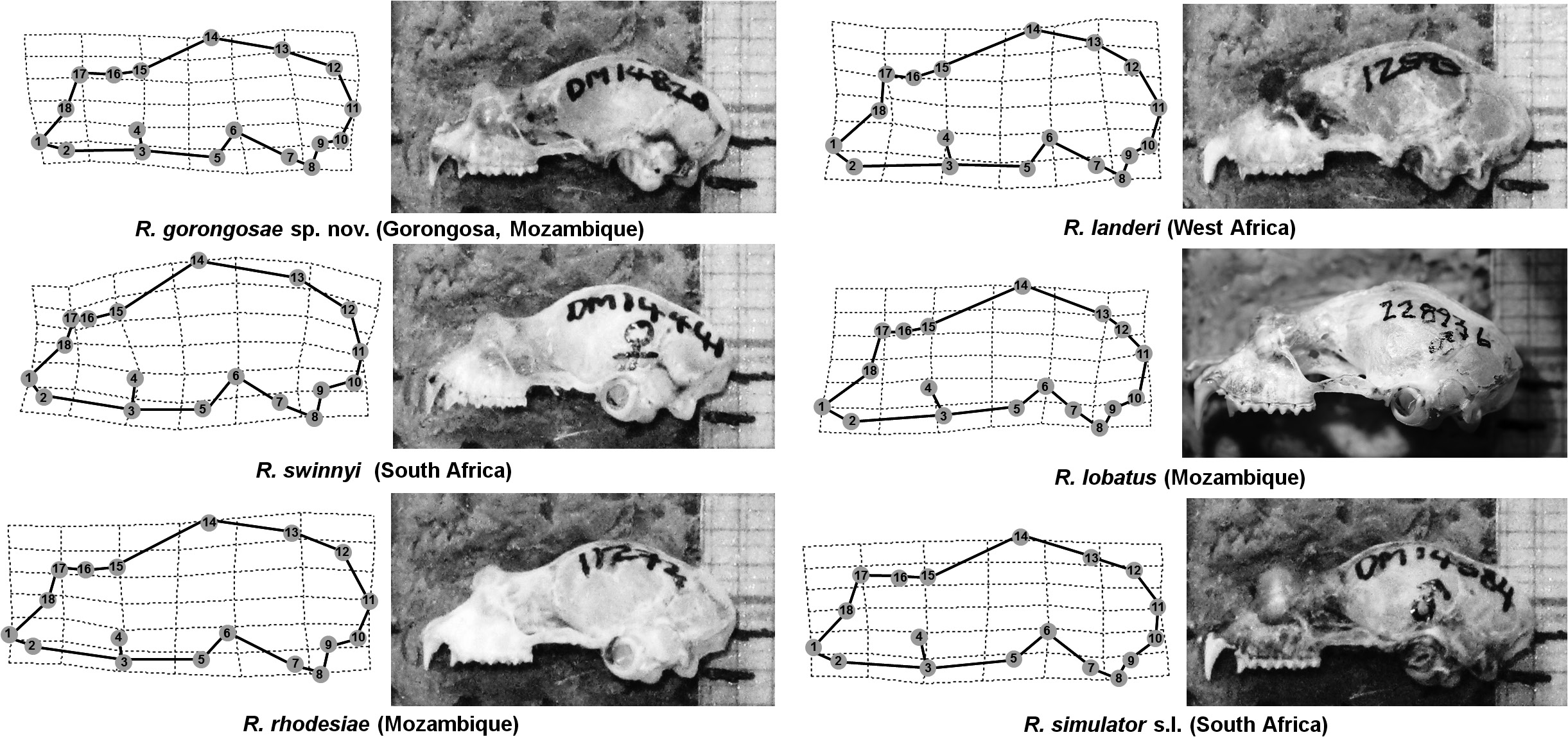
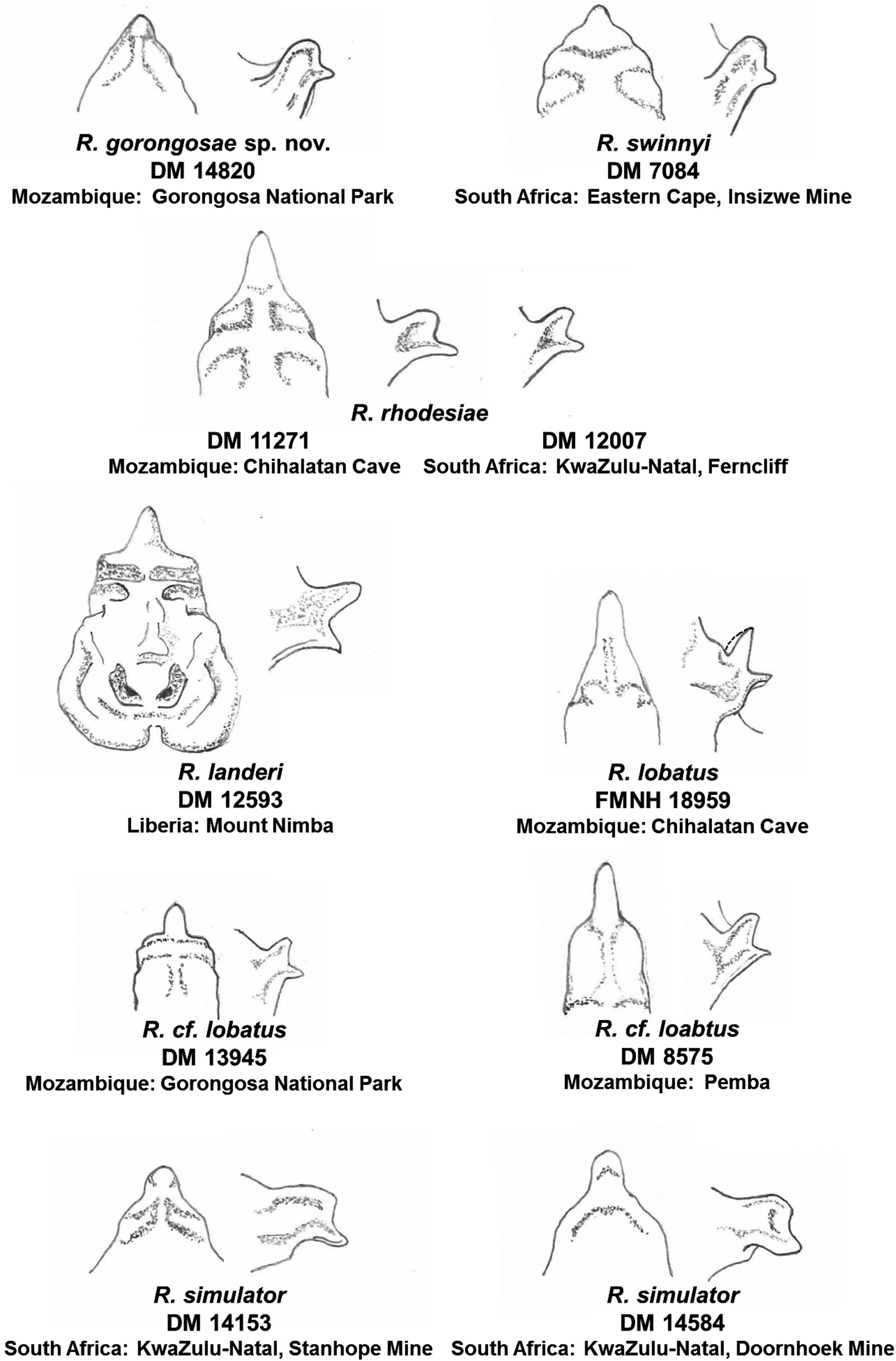
Etymology: The Latin word lobatus means lobed, perhaps referring to the general shape of the noseleaf ( Fig. 13 View Figure 13 ).
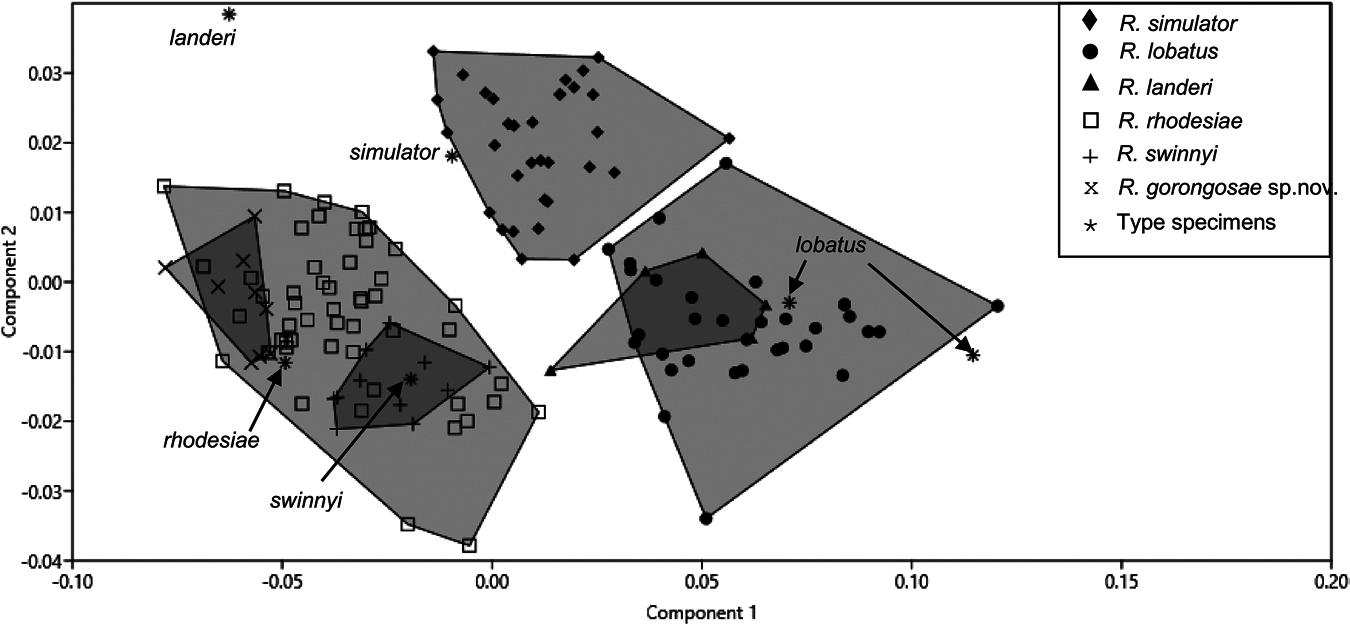
Re-diagnosis and comparisons: This taxon is clearly phylogenetically distinct on Cytb gene sequences (see also: Dool et al., 2016) from topotypic landeri from West Africa and seems to be more closely affiliated with the capensis group of Csorba et al. (2003), i.e. R. capensis , R. denti , R. simulator and R. swinnyi ( Fig. 2 View Figure 2 ). Specimens from Chihalatan and Malashane (N = 4) had a mean echolocation call peak frequency of 106.8 ± 0.4 kHz ( Table 2 View Table 2 ), close to the 107 kHz generally reported for southern Africa ‘ landeri ’ ( Monadjem et al., 2010). This contrasts with a mean maximum frequency of 104.3 ± 0.42 kHz (N = 2) recorded for topotypic West African (Liberian) R. landeri , making a peak frequency of c.107 kHz a possible diagnostic criterion for R. lobatus . However, a wider range (102–108 kHz) and slightly lower mean frequency of 105 kHz was reported for 20 individuals at Gorongosa ( Table 2 View Table 2 ). This variation could be indicative of further undetected cryptic speciation in this taxon, as also indicated by the very wide variation in baculum and noseleaf characters, discussed in the following paragraph.
Although not easily distinguished on body or skull size ( Table 2 View Table 2 ; Fig. 4 View Figure 4 ), this taxon differs from topotypic R. landeri by displaying a prognathic rostrum (LMs 1–3), a shortened braincase that extends posteriorly (LMs 10–14) and a broader nasal inflation (LMs 16–18) ( Figs 5 View Figure 5 , 6 View Figure 6 ). Specimens of cf. lobatus from Pemba (Mozambique) had a shorter foramen magnum (LMs 9–10), a narrower jugal process (LM 3–4) and a slightly broader nasal inflation (LMs 16–18), than other Mozambique animals. The same individual from Pemba possessed a distinctive robust baculum ( Fig. 8 View Figure 8 ) and noseleaf ( Fig. 9 View Figure 9 ) having a markedly hastate lancet and pronounced posterior lobe of the connecting process. However, pending molecular data, we here refer them to lobatus incertae sedis. Although genetically clearly assigned to lobatus , individuals from Gorongosa National Park are somewhat smaller in external (but not cranial) measurements ( Table 2 View Table 2 ), and also display unique noseleaf and bacular morphologies. The noseleaf of a Gorongosa individual has a small hastate lancet and bluntly pointed tip, and a relatively small and more erect connecting process with semi-symmetrical lobe ( Figs 9 View Figure 9 , 13 View Figure 13 ), while its baculum is highly divergent from all other forms considered in our study ( Fig. 8 View Figure 8 ), having a remarkable short baculum with wide shaft and very small base compared with landeri from West Africa, as well as cf. lobatus from Malashane and Chihalatan Caves. Given the clear evidence for close genetic identity in our study between Malashane and Chihalatan Caves and Gorongosa National Park for animals referred to R. lobatus , we provisionally regard this variation in noseleaf and baculum structure to represent polymorphic traits in this species. It is possible that these morphological types represent good species between which introgression has occurred. Further analysis with additional nuclear sequences is necessary to test this hypothesis. There is also the possibility that these could represent subadults with bacula that are not fully ossified, as detailed in R. adami described by Kock et al. (2000).
Male R. lobatus from Chihalatan and Malashane Caves, as well as Gorongosa National Park, typically possessed a dark brown apical tuft of stiff hairs characteristic of R. landeri ( Fig. 14 View Figure 14 ; Csorba et al., 2003; Monadjem et al., 2010). This convergent character could be one of the contributing factors that have led to the historical misidentification of the species with R. landeri .
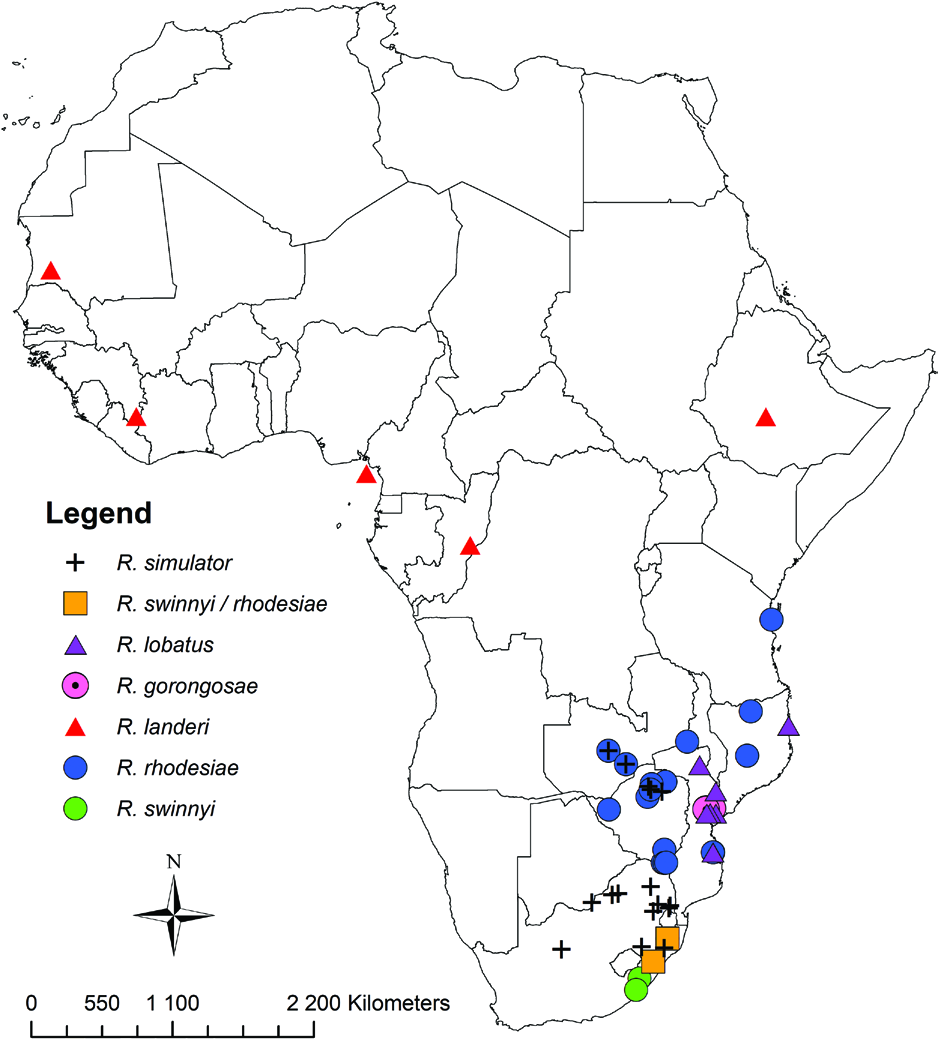
Distribution and biology: As anticipated by Monadjem et al. (2010), who considered that the southern (and possibly eastern) African savannah-occurring lobatus might prove to be distinct from the West African forest-occurring landeri , we here refer all southern African specimens to lobatus ( Fig. 11 View Figure 11 ) [see appendix of Monadjem et al. (2010) for a full list of localities]. We agree with Monadjem et al. (2010) that R. angolensis Seabra, 1989 from western Angola may merit specific status. Nevertheless, in the absence of the type series that was destroyed in the Lisbon fire of 1978, resolution of its status must await a detailed revision, including molecular evidence with designation of new type material. Likewise, the status of east and central African populations must await further studies, including molecular data. Until further evidence becomes available, we suggest that it is prudent to use the name lobatus for all savannah populations in southern and east Africa.





| FMNH |
USA, Illinois, Chicago, Field Museum of Natural History (also used by Finnish Museum of Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |