
Pseudoscopelus scriptus Lütken 1892
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2710.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/852E9C20-FFFF-FFFA-FF3C-FE7D4FD16458 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudoscopelus scriptus Lütken 1892 |
| status |
|
Pseudoscopelus scriptus Lütken 1892 View in CoL
Figures 4 A–J View FIGURE 4 ; 5 A View FIGURE 5 , 6 A View FIGURE 6 , 7 A View FIGURE 7 , 8 A View FIGURE 8 , 28 A; Table 3.
Pseudoscopelus scriptus Lütken 1892: 284–286 View in CoL , plate 1, figures 4–5 [type locality western Central Atlantic, Mouth of “Gamle Bahama Kanalen” (Old Bahamas Channel), holotype ZMUC P. 65171, 70.0 mm]; Goode and Bean 1895: 292–293, plates 76, figure 286 [western North Atlantic]; Parr 1933: 40–41, figure 17 [western North Atlantic]; Norman 1966: 366–367 [key to genus and species list]; Lavenberg 1974: 274–286, figures 51, 54, 55 [western North Atlantic, central Atlantic and Gulf of Mexico]; Uyeno et al. 1983: 404 [western Central Atlantic, off Suriname; questionable]; Bañón et al. 1999 [western North Atlantic, questionable]; Moore et al. 2003: 227 [western North Atlantic]; Prokofiev and Kukuev 2006c [key to species, questionable]; Sptiz et al. 2007: 2–3 [western North Atlantic].
Pseudoscopelus stellatus Beebe 1932: 76–79 View in CoL , figure 18 [type locality Bermuda]; Lavenberg 1974: 303–314, figures 59– 60 [Atlantic and Indian Oceans]; Prokofiev and Kukuev 2005 [questionable]; Prokofiev and Kukuev 2006b: 231, figure 6 A, B; 2008: 122-126, figures 178–179 [redescription of holotype and compilation of Lavenberg 1974]; Sptiz et al. 2007 [mentioned]. NEW SYNONYM.
Pseudoscopelus sagamianus View in CoL (non Tanaka 1908) Prokofiev and Kukuev 2006a: 221–225 [in part, western North Atlantic]; Prokofiev and Kukuev 2008: 112-121 [in part]; Prokofiev 2009: 10–22 [in part, specimens listed for the North Atlantic].
Diagnosis. A species of the Pseudoscopelus scriptus species group, which can be distinguished within the group by a single characteristic: saf not extending anterior to the level of anus (vs. saf extending anteriorly to level of or beyond anus). It can be further distinguished from the other species by a combination of characteristics: mesial series of premaxillary teeth in three to four rows (vs. mesial series of premaxilla in single row P. obtusifrons and P. pierbartus ); mesial series of teeth in rows of three to four teeth gradually increasing of size from lateral to mesial (vs. mesial series arranged in rows of two to three teeth, with internal row much larger than other teeth in P. cordilluminatus ); total vertebrae 35–36, precaudal 18–19 (vs. 31, 14 in P. cephalus ); and pigmentation in the internal part of mouth exclusive to a dusky area in the skin covering regions with teeth on premaxilla and dentary (vs. internal area of mouth and gill arches black in P. sagamianus ).
Description. Middle-sized species, largest specimen examined 203.0 mm SL. Morphometric data summarized in Table 3. General body shape as described for genus with diagnostic characteristics of species and species group. Snout moderately blunt; anterior naris lateral in snout.
First dorsal-fin rays VII (1), VIII (7*), IX (4); second dorsal-fin rays ii+19 (1), ii+20 (3), ii+21 (7*), ii+22 (1); anal-fin rays iii+19 (3), iii+20 (5*), iii+21 (3), iii+22 (1); pectoral-fin rays 12 (2), 13 (7*), 14 (3); pelvicfin rays I+5 (12*); caudal-fin rays i+7+8+i (11*), i+8+8+i (1). Branchiostegal rays 7 (12*). Pre-caudal vertebrae 18 (3), 19 (5); total vertebrae 35 (1), 36 (7).
Lateral line complete; lateral-line pores 73 (1), 74 (1), 75 (3*), 77 (4), 78 (2). Pores in temporal canal 2 (12*); supratemporal canal 3 (12*); otic canal 2 (12*); supraorbital canal 5 (11*), 6 (1); supranasal pore 2 (10*), 1 (2); epiphyseal branch 2 (5), 3 (5*); infraorbital canal 11 (11*), 12 (1); preopercular canal 5 (13*); mandibular canal 6 (13*); fifth pore of mandibular canal 2 (12*).
Dentition. Enlarged teeth on premaxilla, dentary and palatine. Premaxilla and dentary illustrated in Figure 5 A View FIGURE 5 . Premaxilla moderately wide, widest point of premaxillary body 15–17 % in premaxillary length.
Premaxillary teeth on head, neck, body and caudal process. Lateral series in single longitudinal row, along lateral edge of premaxillary head, neck, body and caudal process; teeth conical, slightly curved. Canine and fang on ventral shelf of premaxillary head. Middle and mesial series of premaxilla on ventral shelf of body. Middle series in two, irregular, longitudinal row; teeth needle-like, straight, gradually increasing in size from lateral to medial. Mesial series in transverse rows, each row with three to four teeth; teeth needle-like, slightly curved, gradually increasing in size from medial to mesial.
Dentary teeth in lateral and mesial series. Lateral series along lateral shelf of dentary, in single, longitudinal row, extending from symphysis to posterior tip; teeth conical, slightly curved. Mesial series in transverse rows of one to four teeth; teeth straight, needle-like, gradually increasing in size from lateral to mesial. Palatine teeth 3 (1), 4 (2), 6 (3), 7 (5*), 8 (1); in single, longitudinal row.
Teeth on infrapharyngobranchials and fifth ceratobranchial, conical, curved. Teeth on second basibranchial 5 (2), 6 (1), 7 (1), 9 (2), 10 (4), 11 (2*), conical, in V -shaped or single, irregular row. Teeth absent on basihyal and other basibranchials. Gill rakers on first epibranchial 0 (12*); first ceratobranchial 6 (1), 7 (3), 8 (3), 9 (1), 10 (1), 11 (3*); first hypobranchial 2 (2), 4 (3), 5 (2), 6 (2), 7 (3*). Gill rakers absent on other elements.
Luminescent organs. Luminescent organs present as discrete photophores on head and body ( Fig. 6 A View FIGURE 6 ). Photophores on head: apf, dnf, inof 1–2, lpf, opf, and pof absent; mxf elongated, in single row anteriorly and two posteriorly, parallel to maxilla, from level of posterior edge of eye to angle between preopercle and dentary; vnf usually absent, but present in some specimens; ppf in small patch on ventral edge of interopercle; amf in two to three rows, medial to mandibular canal, from second pore to halfway between third and fourth pores; pmf in one to three rows, lateral to mandibular canal, from halfway between fourth and fifth pores to halfway between fifth and sixth pores of mandibular canal.
Photophores on body: lvf, rtf, scf, spf, and svf absent; pf in single row extending to half pectoral fin, on ventral ray; paf continuous with pf, at pectoral-fin axil; vf in single row, along mesial pelvic-fin ray; vaf continuous with vf and trf, extending over base of pelvic rays three to five; if and prvf continuous, in irregular two to three rows, from isthmus to anterior part of pelvic girdle; ptvf in two rows, from posterior half of pelvic fin to close to anus; trf laterally in single row, with medial circular group of photophores; saf in two rows, with smaller photophores closely spaced and ventral, and larger photophores widely spaced and dorsal, heartshaped, from far anterior to anus, to almost halfway between pelvic and anal fins, and connected posteriorly; prcf in posterior half of peduncle, oval-shaped or three-pronged, medial prong extending over anteriormost lower procurrent rays.
Color. Most specimens examined faded to light brown or bleached white, including holotype. Body uniformly black or dark brown, except for triangular area on epiphyseal branch. Pectoral and pelvic fins hyaline; anal, first and second dorsal fins with melanophores uniformly distributed over rays, more concentrated on fin base; and caudal fin pigmented at base. Internal area of mouth and gill arches pale.
Distribution. In the Western Atlantic from the United States to Brazil, continental shelf and Islands of the Caribbean in the west to Central Atlantic; from 69º N to 20º S, 39º W to 23º W ( Fig. 7 View FIGURE 7 ). A juvenile specimen was tentatively identified from off Nigeria (5º56’ N, 4º03’ E), possibly extending the species range to the Eastern Atlantic.
Bathymetric distribution. Meso- to bathypelagic, from 250 to 1370 m (mean 786 m); juveniles to about 30.0 mm are known from 95 to 590 m.
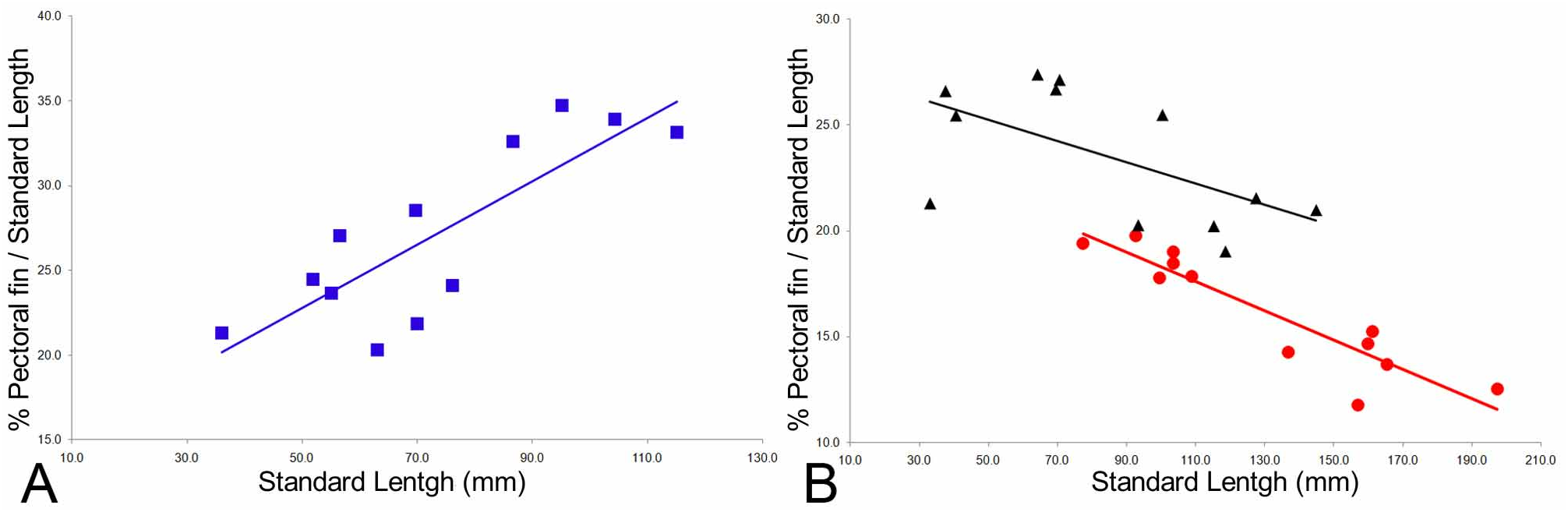
Identification of juveniles. Juveniles of Pseudoscopelus scriptus are not distinguishable from those of P. pierbartus by dentition pattern, but were tentatively identified by the shape of saf, not extending to level of anus. Pseudoscopelus scriptus has positive allometric grow of the pectoral fin (P=0.001, Fig. 8 A View FIGURE 8 ).
Remarks. Lavenberg (1974) tentatively distinguished Pseudoscopelus scriptus from P. sagamianus based on three characteristics: anterior photophores of mxf below ventral margin of orbit (vs. anterior photophores below middle of eye); presence of vnf (vs. vnf absent); and single aperture in supranasal pore (vs. two in P. sagamianus ). None of Lavenberg's characters seems to be useful because of individual variation. In P. sagamianus , the anterior photophores of mxf can also extend anteriorly to below mid-eye; vnf is variable, it is absent in most specimens of P. scriptus , including the holotype, but present in a few specimens (e.g., MCZ 101790; the same is true for P. sagamianus , see species account); also, most specimens of P. scriptus , including the holotype, have two supranasal pores.
Prokofiev and Kukuev (2006 a, 2006c, 2008) and Prokofiev (2009) were unable to correctly identify this species. The diagnosis of Pseudoscopelus scriptus provided by Prokofiev and Kukuev (2006a: 218–221) is imprecise and erroneous, despite their statement that “the identity of the aforementioned fish [ P. scriptus ] is not in doubt” (p.221). Characteristics of dentition described and illustrated as “manifestations of individual variation” (p. 221, Figs. 2 F, G View FIGURE 2 ), actually reflect the pattern of two different species, P. pierbartus and P. cordilluminatus . Prokofiev and Kukuev (2008: 102–112, Figs. 158, 159) perpetuated the mistake. Moreover, several specimens listed as P. sagamianus for the North Atlantic ( Prokofiev and Kukuev 2006 a, 2008; Prokofiev 2009), actually belong to P. scriptus (see discussion for P. sagamianus ).
Sptiz et al. (2007) provided insufficient diagnostic data for Pseudoscopelus scriptus , missing the characteristics of dentition that actually distinguish it from P. pierbartus .
The status of Pseudoscopelus stellatus . Pseudoscopelus stellatus was described based on five larval specimens between 6.5 mm and 20.3 mm, from Bermuda, western North Atlantic – a brief description of the specimens is given by Melo et al. (2007). Eschmeyer (1998) considered the types to be lost, but later Eschmeyer (2007) listed a lot from the AMNH and three others from CAS as holotype and paratypes, respectively.
Lavenberg (1974) considered P. stellatus as valid and listed several larvae under the species account. Prokofiev and Kukuev (2006c) considered this species valid but did examine neither the types nor additional material from the region. Later, Prokofiev and Kukuev (2008) followed the original description to consider it as valid and provided illustrations which do not accurately reflect the photophore pattern of the holotype. Melo et al. (2007) were not able to determine its correct identification and preferred to leave it as species inquirenda.
Beebe (1932) was the first author to notice the existence of photophores in Pseudoscopelus , and illustrated and described the photophore groups in the holotype of P. stellatus . The specimen, however, is a juvenile and after the examination of more specimens, it becomes clear that not all photophore groups are fully developed in the specimen used. Moreover, only the holotype (USNM 170941) has photophores and the other specimens used for the description lack the photophores and are assigned to P. aphos ( Figs. 4 D–G View FIGURE 4 ).
Seven species are known from around the Bermuda area: Pseudoscopelus altipinnis , P. aphos , P. astronesthidens , P. scutatus , P. pierbartus , P. scriptus , and P. obtusifrons . Besides the poor condition of the holotype ( Figs. 4 D, H–J View FIGURE 4 ), a close examination of it revealed some details that were not described by Beebe (1932) but are useful to establish the correct identity of Pseudoscopelus stellatus : photophores present (vs. absent in P. aphos ); saf (=circum-anal of Beebe 1932) not interrupted anteriorly (vs. interrupted anteriorly in P. astronesthidens ); if and prvf continuous and separate from ptvf (vs. if and prvf separate in P. altipinnis and P. scutatus , and prvf and ptvf continuous in the latter). The mxf is in a small group but, different from what was illustrated, it is not oval; instead, it is arranged in a straight line ( Figs. 4 J View FIGURE 4 ), a condition that is only found in the P. scriptus species group.
The most remarkable characteristic of the holotype is the shape of saf, which does not extend anteriorly to the level of anus, and is connected posteriorly behind the last anal-fin ray (vs. saf extending to the level of, or slightly beyond anus and not connected posteriorly in Pseudoscopelus obtusifrons and P. pierbartus ). That shape of saf only fits with P. scriptus . Although some of the diagnostic photophores for that species are not present (trf, ppf, prcf, pf, vf, and vaf), this can be understood because the specimen a juvenile and some of the groups are not yet developed. If an adult specimen missing those photophores becomes available, P. stellatus would perhaps be valid, but the evidence for now is that P. stellatus is a junior synonym of P. scriptus .
Other three specimens used for the description (CAS- SU 66496 and CAS-SU 66497), which were considered to be paratypes by Eschmeyer (2007) lack photophores. In other species such as P. scriptus , juveniles of the same size have some groups of photophores present on ventral part of body and head. Therefore, those specimens are identified herein as P. aphos .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pseudoscopelus scriptus Lütken 1892
| Melo, Marcelo R. S. 2019 |
Pseudoscopelus sagamianus
| Prokofiev, A. M. 2009: 10 |
| Prokofiev, A. M. & Kukuev, E. I. 2008: 112 |
| Prokofiev, A. M. & Kukuev, E. I. 2006: 221 |
Pseudoscopelus stellatus
| Prokofiev, A. M. & Kukuev, E. I. 2006: 231 |
| Lavenberg, R. J. 1974: 303 |
| Beebe, W. 1932: 79 |
Pseudoscopelus scriptus Lütken 1892: 284–286
| Moore, J. A. & Hartel, K. E. & Craddock, J. E. & Galbraith, J. K. 2003: 227 |
| Uyeno, T. & Matsuura, K. & Fujii, E. 1983: 404 |
| Lavenberg, R. J. 1974: 274 |
| Norman, H. W. 1966: 366 |
| Parr, A. E. 1933: 40 |
| Goode, G. B. & Bean, T. H. 1895: 292 |
| Lutken, C. F. 1892: 286 |