
Chilerium zapallar, (Chamberlin, 1955)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4027.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:4218E1CD-9016-47C3-A1DA-291F9C53661F |
|
DOI |
https://doi.org/10.5281/zenodo.5667414 |
|
persistent identifier |
https://treatment.plazi.org/id/875F87C1-FFA5-3C31-FF11-ECF7340BFCEB |
|
treatment provided by |
Plazi |
|
scientific name |
Chilerium zapallar |
| status |
|
Plateurytion zapallar ( Chamberlin, 1955)
( Figs. 54–109 View FIGURES 54 – 58 View FIGURES 59 – 66 View FIGURES 67 – 74 View FIGURES 75 – 80 View FIGURES 81 – 88 View FIGURES 89 – 94 View FIGURES 95 – 98 View FIGURES 99 – 104 View FIGURES 105 – 109 )
Chilerium zapallar Chamberlin, 1955: 24 , 25.
Eurytion zapallar: Foddai, Pereira & Minelli, 2000:75 View in CoL , 185. Eurytion zapallar: Pereira, 2006:167 View in CoL (in key).
Plateurytion zapallar View in CoL : Bonato, Pereira & Minelli, 2007:6; Pereira, 2008:56.
Diagnosis. A species of Plateurytion characterized by having one cluster of coxal organs in each coxopleuron of the ultimate leg-bearing segment. Of the other South American species currently included in the genus, only the present species and P. mundus ( Chamberlin, 1955) share the same character. Plateurytion zapallar can be confidently differentiated from P. mundus by means of the characters used in the preceding diagnosis of the latter.
Other morphological traits included in Table 1 View TABLE 1 differentiate P. zapallar from P. mundus .
Remarks. P. zapallar can be separated from the other South American members of Plateurytion using the identification key below.
Type material examined. CHILE: Aconcagua: Zapallar , 27 November, 1950 (collector?), two syntypes here designated as lectotype ♂ with 49 l. -b.s., (b.l. 28 mm?) (head capsule with mandibles, dissected first and second maxillae, leg-bearing segments 42-49 and postpedal segments, in an original permanent slide; 26 l. -b.s. of anterior region of the body, in two parts of 5 and 21 each, in alcohol); paralectotype ♀, subadult, with 49 l. -b.s., b.l. 17 mm (undissected, in alcohol). Both specimens labeled as Chilerium zapallar Chamberlin ( CAS Entomology type Nº 9174).
Remarks. Chamberlin (1955) stated that the syntypical series comprised three specimens (all with 49 legbearing segments), but actually comprises two (apparently one of these is now missing); the author gives 28 mm as body length for the species, because the trunk of the lectotype is incomplete, it is not possible to determine if this length corresponds to it (or to the presumptive missing specimen of unknown sex). Lectotype male with forcipular segment and 15 leg-bearing segments of posterior region of the trunk, missing. Paralectotype female complete, showing ova and spermathecae without spermatozoa (located at level of leg-bearing segment 48).
Depository of types. CAS.
Other material examined. CHILE: Region IV (Coquimbo region): Elqui province: 11 km N of Los Vilos, 11 November 1987, E. Maury coll.: 1 ♀ with 47 l. -b.s., b.l. 26 mm (MACN-My 25).
Region V (Valparaíso region). Petorca province: Quebrada Huaquén, Pichicuy, 7 January 1984, E. Maury coll.: 1 ♂ with 47 l. -b.s., b.l. 22 mm (MACN-My 26). Quillota province: Parque Nacional La Campana, Palmas de Ocoa, 8–5 November 1987, E. Maury coll.: 1 ♀ with 53 l. -b.s., b.l. 28 mm (MACN-My 27); same locality and collector, 27–28 October 1988, 1 ♂ with 51 l. -b.s., b.l. 23 mm (MACN-My 28); same locality, 32º 57´40.4´´ S, 71º 03´34.0´´ W, 770 m a.s.l., 18 February, 2005, M. J. Ramírez & F. Labarque coll.: 1 ♂ with 51 l. -b.s., b.l. 25 mm (MACN-My 29). Valparaíso province: Quebrada el Tigre, Cachagua, 8 November 1988, E. Maury coll.: 1 ♂ with 47 l. -b.s., b.l. 27 mm (MACN-My 30).
Redescription. Male (specimen MACN-My 29). Fifty-one leg-bearing segments, body length 25 mm, maximum body width 1.32 mm. Maximum width of cephalic plate 0.73 mm, length of cephalic plate 1.12 mm, maximum width of forcipular coxosternite 1.00 mm. Ground color (of preserved specimen in alcohol) yellowish, forcipular segment darker (pale ochreous).
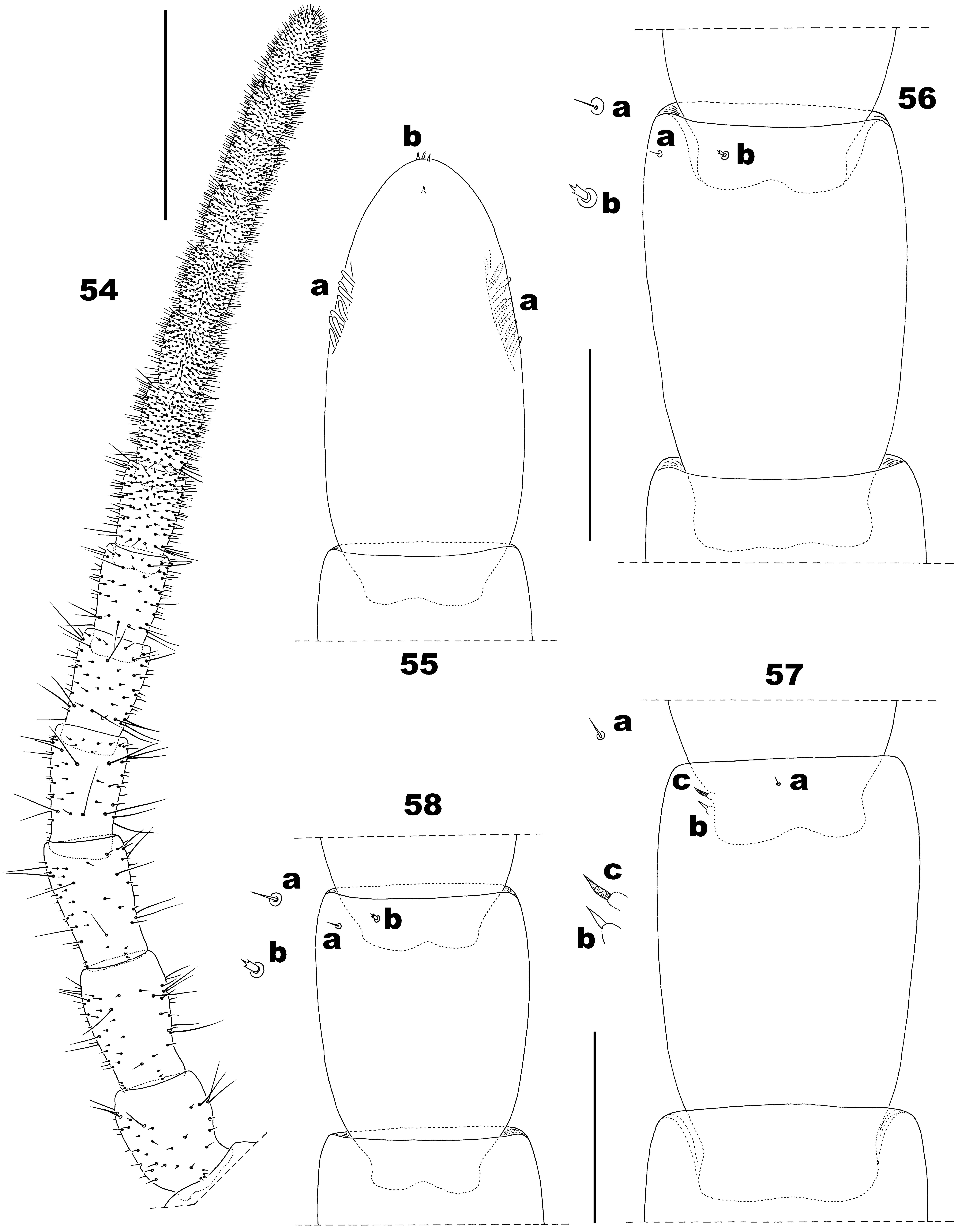
Antennae. Relatively short, ca. 2.64 times as long as the cephalic plate, distally attenuate ( Fig. 54 View FIGURES 54 – 58 ). Ratio of width of a.a. II/width of a.a. XIV ca. 2.04: 1; all a.a. longer than wide. Ventral chaetotaxy: setae on a.a. I–VI of various lengths and relatively few in number, those of a.a. VII–XIV progressively shorter and more numerous towards the tip of the appendage ( Fig. 54 View FIGURES 54 – 58 ). Dorsal chaetotaxy: setae on a.a. I–VI similar to the ventral side, setae on remaining a.a. a little longer and slightly less numerous. A.a. XIV with ca. 13 claviform sensilla on the external margin and ca. 9 on the internal margin ( Fig. 55 View FIGURES 54 – 58 : a); distal end of this a.a. with ca. 4–5 very small hyaline specialized sensilla not split apically ( Fig. 55 View FIGURES 54 – 58 : b). Ventral and dorsal surface of a.a. II, V, IX and XIII with very small specialized sensilla. On the ventral side these sensilla are placed in the internal latero-apical area and are represented by two different types: a and b. Type a sensilla are very thin and not split apically ( Fig. 58 View FIGURES 54 – 58 : a); type b sensilla ( Fig. 58 View FIGURES 54 – 58 : b) are very similar to those on the apex of a.a. XIV but having two diminutive apical branches. Specialized sensilla on dorsal side restricted to a middle and external latero-apical areas and are represented by three different types: type a, similar to type a of ventral side ( Fig. 59 View FIGURES 59 – 66 : a), type b sensilla not split apically and similar to those on the apex of a.a. XIV ( Fig. 59 View FIGURES 59 – 66 : b), and type c sensilla similar in shape to the latter, a little larger, not divided apically and slightly darker (pale brownish-ochreous in color) ( Fig. 59 View FIGURES 59 – 66 : c). Number and distribution of specialized sensilla on ventral and dorsal sides of a.a. II, V, IX and XIII, as in Table 3.
Ventral Dorsal Figs. a b a b c
II – 1 – 1 –
V 1 1 1 1 –
IX 1 1 1 1 1 56, 57
XIII 1 1 1 3 1 58, 59 Cephalic plate. Distinctly longer than wide (length/width ratio ca. 1.48: 1). Middle part of lateral margins convergent towards the posterior region; anterior margin convex at middle, slightly concave at level of the bases of the antennae; posterior margin nearly straight; anterior and posterior sides, curved. Shape and chaetotaxy as in Fig. 60 View FIGURES 59 – 66 .
Clypeus. With 1 + 1 setae adjacent to the anterior margin of the clypeal area, two setae located on it ( Fig. 62 View FIGURES 59 – 66 ), posterior to the latter 2 + 2 setae distributed at both sides of the middle line ( Fig. 61 View FIGURES 59 – 66 ). Clypeal area well developed with surface minutely punctuate or granulate, not areolate ( Fig. 62 View FIGURES 59 – 66 ).
Labrum. Mid-piece well developed, subtriangular in shape, slightly pigmented and provided with 13 hyaline and thin filaments ( Figs. 63, 64 View FIGURES 59 – 66 ). Side pieces with 17 + 17 hyaline filaments of different sizes ( Fig. 63 View FIGURES 59 – 66 ).
Mandible. With shape as in Fig. 65 View FIGURES 59 – 66 , pectinate lamella with ca. 22 hyaline teeth.
First maxillae. Coxosternite and telopodites with very well developed lappets ( Fig. 66 View FIGURES 59 – 66 ). Coxosternite devoid of setae; coxal projections subtriangular, round-tipped and provided with 6 + 6 setae and 2 + 2 very small sensilla ( Fig. 66 View FIGURES 59 – 66 ). Article II of telopodites with 7 + 5 large setae on ventral side ( Fig. 66 View FIGURES 59 – 66 ), and 1 + 1 small sensilla on dorsal side.
Second maxillae. Coxites medially joined through a narrow, hyaline and non-areolate membranous isthmus only and provided with 8 + 8 setae near the internal and antero-internal borders and 2 + 2 setae near the external borders ( Fig. 66 View FIGURES 59 – 66 ). Metameric pores accompanied by a sclerotized rim with shape as in Fig. 66 View FIGURES 59 – 66 : c. Apical claw of telopodites well developed ( Fig. 66 View FIGURES 59 – 66 ). Chaetotaxy of coxosternites and telopodites as in Fig. 66 View FIGURES 59 – 66 .
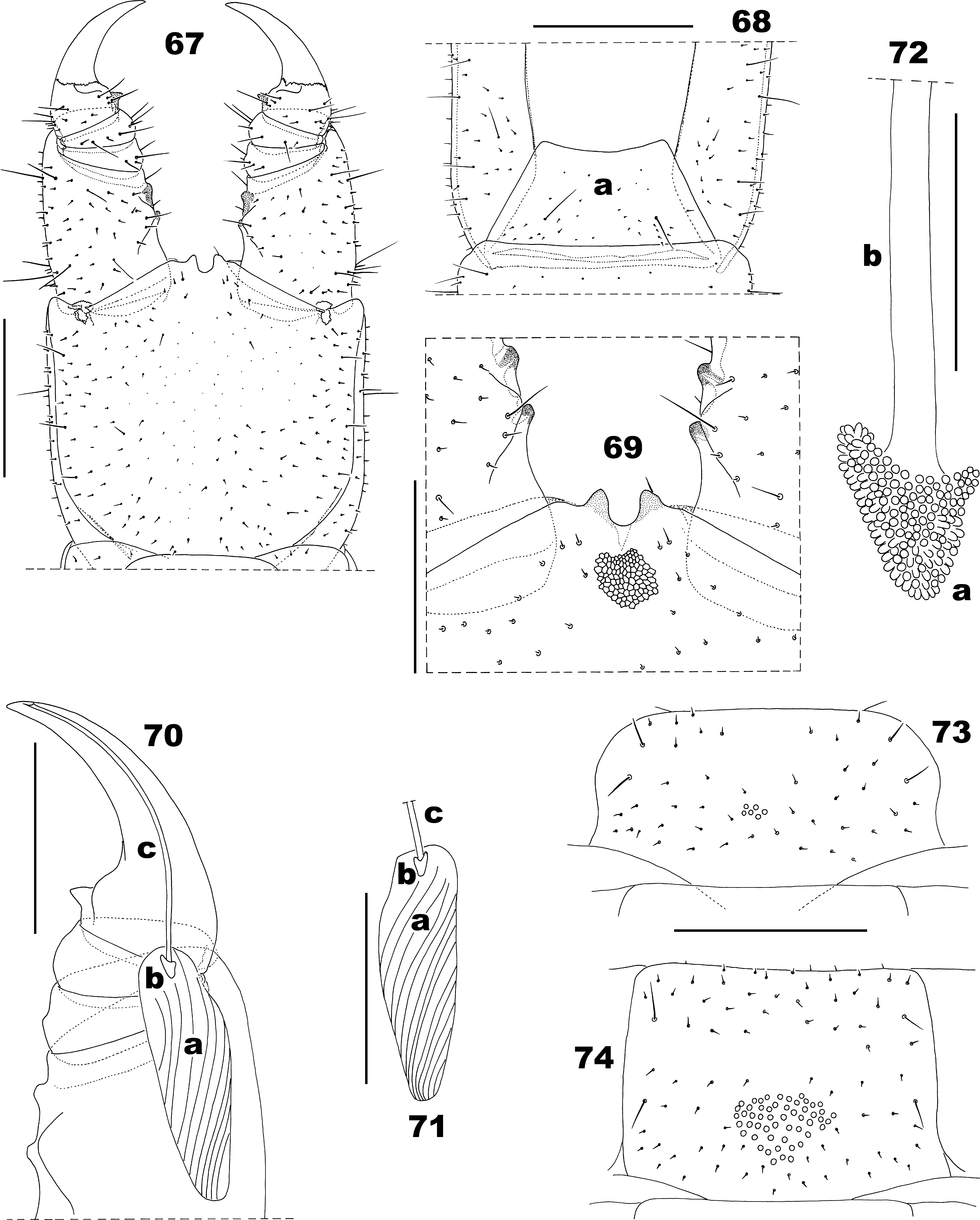
Forcipular segment. When extended the telopodites attaining the middle part of the a.a. II. Forcipular tergite trapeziform ( Fig. 62 View FIGURES 59 – 66 : a), with anterior and posterior margins respectively covered by the cephalic plate and the tergite of the first leg-bearing segment; chaetotaxy represented by 1 + 1 large setae and ca. 17 + 17 very small setae with relative size and distribution as in Fig. 68 View FIGURES 67 – 74 . Coxosternite without chitin-lines, middle part of anterior border conspicuously notched and strongly concave, provided with 1 + 1 slightly pigmented denticles, the left one bearing a dorsal seta; aspect and relative size as in Figs. 67, 69 View FIGURES 67 – 74 . Telopodites: medial edge of trochanteroprefemur with a proximal and a distal tooth, both deeply pigmented ( Figs. 67, 69 View FIGURES 67 – 74 ). Femur and tibia without denticles. Tarsungulum basally with a well developed and deeply pigmented subtriangular tooth ( Figs. 67, 70 View FIGURES 67 – 74 ); ventral medial edge of tarsungulum not serrate ( Figs. 67, 70 View FIGURES 67 – 74 ). Relative size of poison gland as in Fig. 70 View FIGURES 67 – 74 , calyx of poison gland subtriangular in shape ( Figs. 70–72 View FIGURES 67 – 74 ). Chaetotaxy of coxosternites and telopodites as in Figs. 67, 69 View FIGURES 67 – 74 .
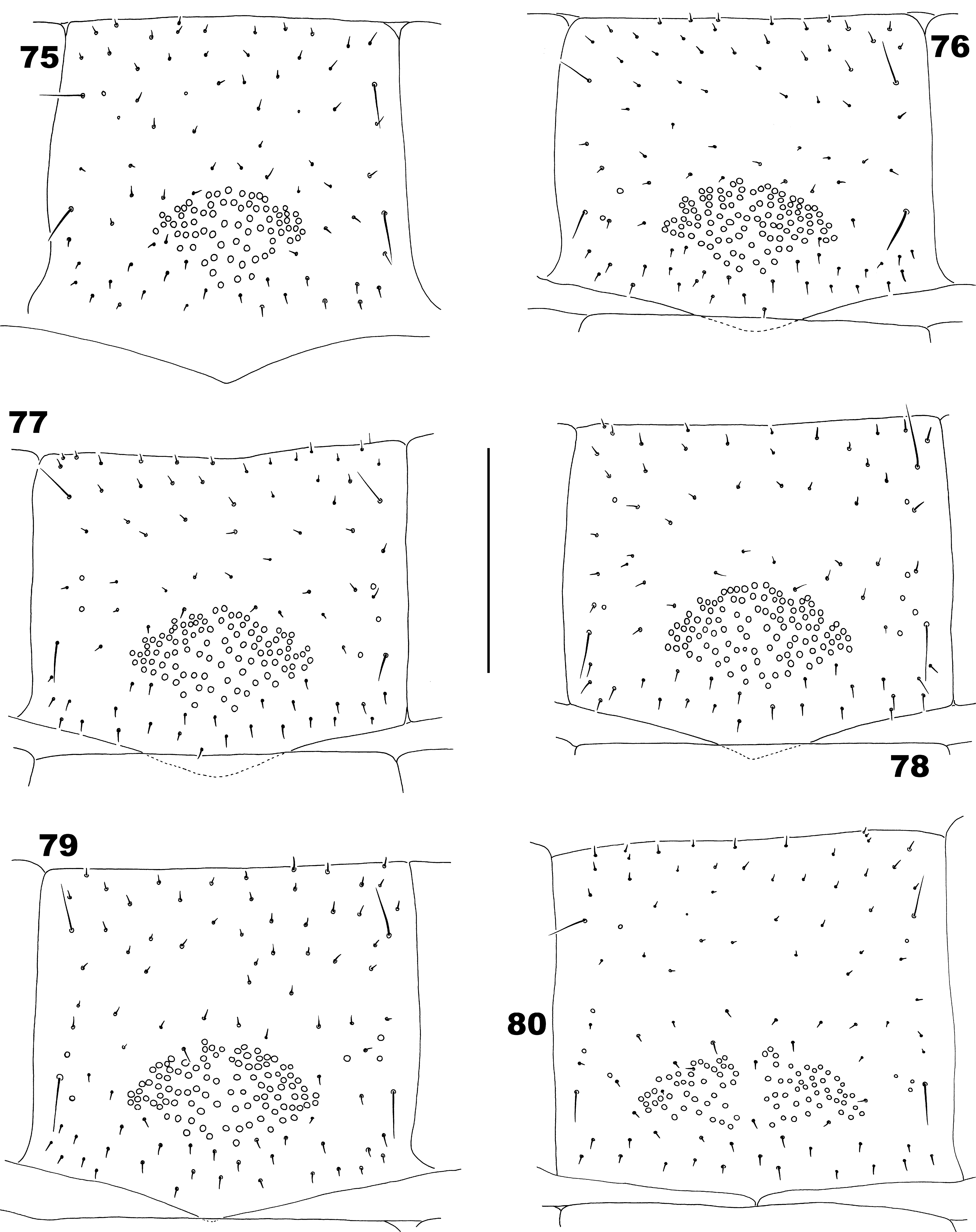
Metasternites of leg-bearing segments 1 to penultimate. With a median shallow longitudinal sulcus all along the body length. Pore-fields present in an uninterrupted series from metasternite 1 to penultimate inclusive; fields undivided on metasternites 1–15 ( Figs. 73–79 View FIGURES 67 – 74 View FIGURES 75 – 80 ), divided in two subsymmetrical areas on metasternites 16–50 ( Figs. 80–85 View FIGURES 75 – 80 View FIGURES 81 – 88 ). Metasternites 4 ( Fig. 75 View FIGURES 75 – 80 ); 6; 7 ( Fig. 76 View FIGURES 75 – 80 ); 8; 9; 10 ( Fig. 77 View FIGURES 75 – 80 ); 11; 12 ( Fig. 78 View FIGURES 75 – 80 ); 13 ( Fig. 79 View FIGURES 75 – 80 ); 14; 15; 16 ( Fig. 80 View FIGURES 75 – 80 ); 17; 18 ( Fig. 81 View FIGURES 81 – 88 ); 19–24; 26; 34; 37–43; 45; 46 ( Fig. 83 View FIGURES 81 – 88 ); and 47 with ca. 1–5 additional dispersed pores near one or both lateral margins. Number of pores on selected metasternites as follows: metasternite 1 (6); 2 (53); 4 (1 + 71 + 0); 7 (2 + 99 + 0); 10 (2 + 97 + 3); 12 (2 + 106 + 5); 13 (2 + 99 + 3); 16 (4 + 41 + 44 + 4); 18 (3 + 31 + 34 + 3); 32 (11 + 11); 46 (1 + 15 + 14 + 0); 49 (10 + 16); 50 (9 + 6). Chaetotaxy of metasternites, shape and relative size of pore-fields as in Figs. 73–85 View FIGURES 67 – 74 View FIGURES 75 – 80 View FIGURES 81 – 88 .
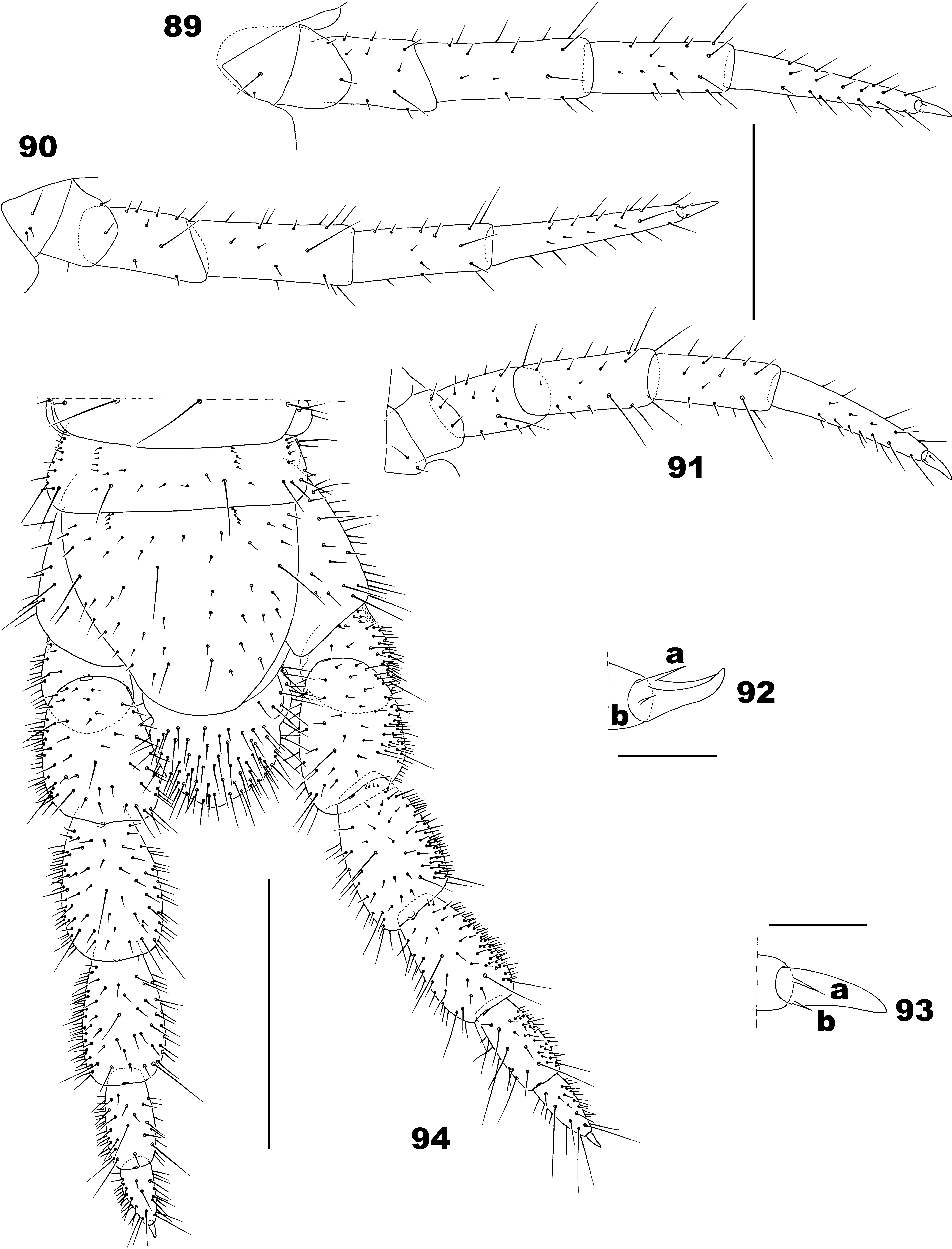
Legs (pair 1 to penultimate). Ratio of length of first pair/length of second pair ca. 0.80: 1 (relative size as in Figs. 86, 87 View FIGURES 81 – 88 ). Chaetotaxy similar throughout the whole body length; distribution, number and relative size of setae as in Figs. 86–91 View FIGURES 81 – 88 View FIGURES 89 – 94 . Claws with two thin and pale accessory spines ventrobasally, the anterior ( Figs. 92, 93 View FIGURES 89 – 94 : a) bigger than the posterior ( Figs. 92, 93 View FIGURES 89 – 94 : b).
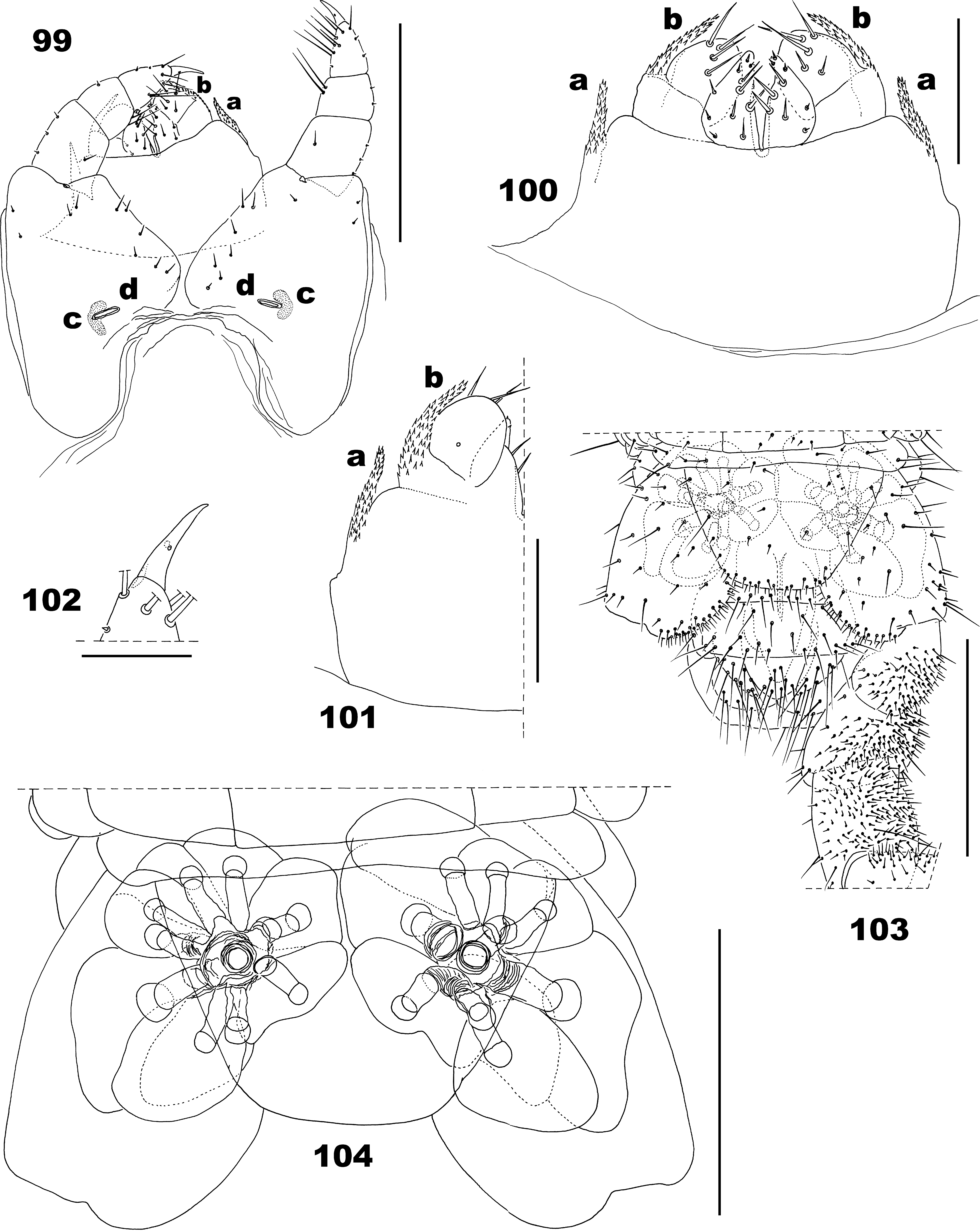
Ultimate leg-bearing segment. Intercalary pleurites absent at both sides of the ultimate pretergite ( Fig. 94 View FIGURES 89 – 94 ); ultimate presternite not divided along the sagittal plane ( Fig. 95 View FIGURES 95 – 98 ). Length/width ratio of metatergite, ca. 0.87: 1; length/width ratio of metasternite, ca. 0.79: 1. Shape and chaetotaxy of metatergite and metasternite as in Figs. 94 View FIGURES 89 – 94 , 95 View FIGURES 95 – 98 . Coxopleura strongly protruding at their distal-internal ventral ends, setae small and numerous on the internal ventral area, the remaining coxopleural surface with much less numerous setae of different lengths ( Figs. 94 View FIGURES 89 – 94 , 95 View FIGURES 95 – 98 ). Each coxopleuron with all coxal organs grouped in a cluster opening on the membrane between coxopleuron and metasternite, partially or totally covered by the latter ( Figs. 95, 96 View FIGURES 95 – 98 ). Right cluster with 11 organs; left cluster with 10 organs; arrangement of organs in each cluster as in Fig. 96 View FIGURES 95 – 98 . Telopodites of ultimate legs composed of six conspicuously inflated articles. Ratio of length of telopodites of ultimate legs/length of metasternite ca. 3.98: 1. Shape and chaetotaxy of ultimate legs as in Figs. 94 View FIGURES 89 – 94 , 95 View FIGURES 95 – 98 . Ultimate pretarsus unguiform, relatively smaller than those of the preceding legs, bearing a single very small and hyaline internal spine ventrobasally.
Postpedal segments. Intermediate tergite with posterior margin strongly convex ( Fig. 94 View FIGURES 89 – 94 ), intermediate sternite with posterior margin nearly straight ( Fig. 95 View FIGURES 95 – 98 ). Posterior margin of first genital sternite concave ( Fig. 95 View FIGURES 95 – 98 ). Gonopods apparently uniarticulate (suture between the presumptive basal and distal articles not evident), bearing ca. 25 setae ( Fig. 97 View FIGURES 95 – 98 ); penis dorsally with 3 + 3 apical setae ( Fig. 98 View FIGURES 95 – 98 ). Anal organs absent.
Female (specimen MACN-My 25). Forty-seven leg-bearing segments, body length 26 mm, maximum body width ca. 1.25 mm. Features similar to those in the male, except for the shape and pilosity of the ultimate legbearing segment and postpedal segments.
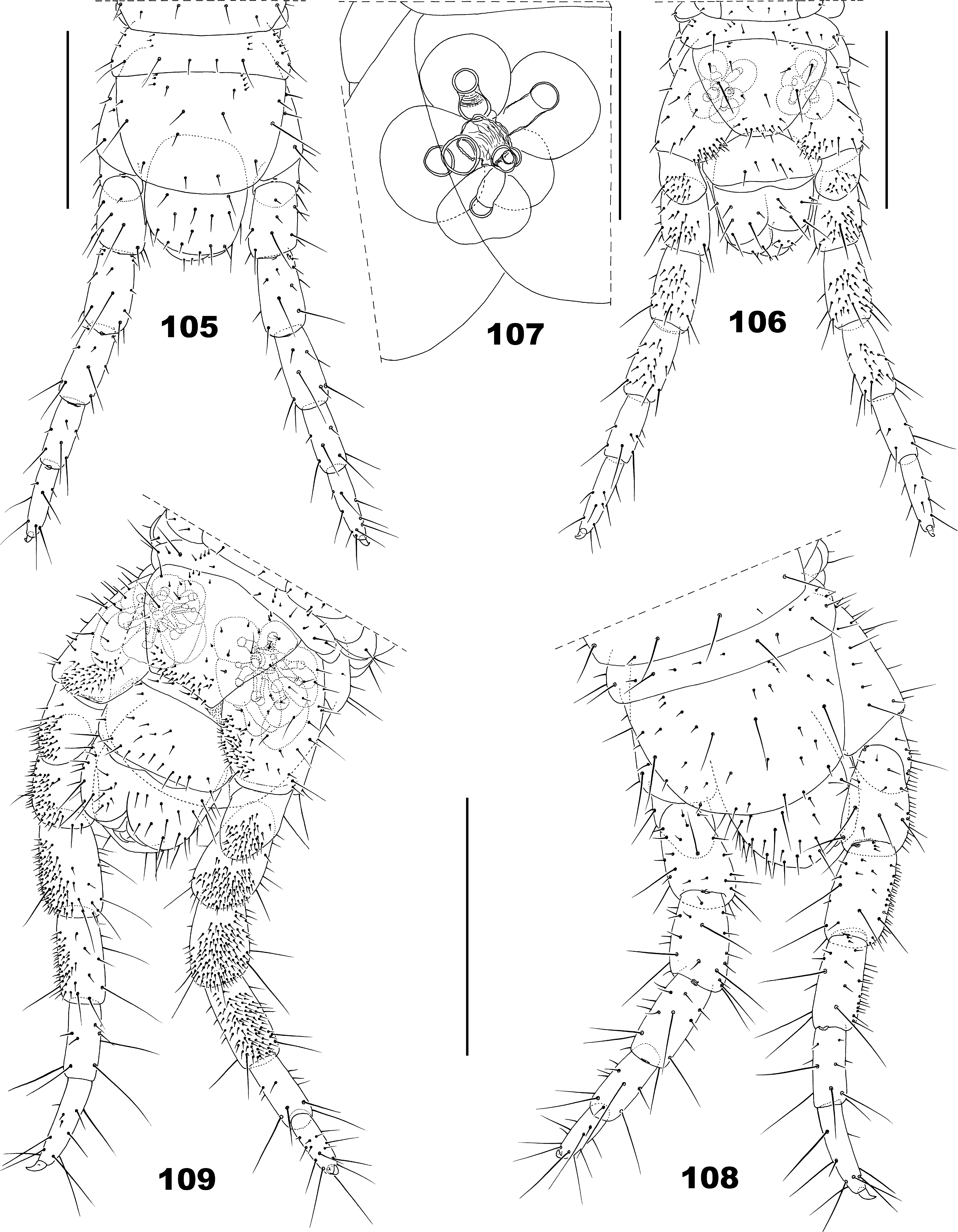
Ultimate leg-bearing segment. Length/width ratio of metatergite 0.65: 1; length/width ratio of metasternite 0.72: 1. Shape and chaetotaxy of metatergite and metasternite as in Figs. 108, 109 View FIGURES 105 – 109 . Coxopleura conspicuously protruding at their distal-internal ventral area, setae small and numerous on the internal ventral area, the remaining coxopleural surface with much less numerous setae of different lengths ( Figs. 108, 109 View FIGURES 105 – 109 ). Right and left cluster of coxal organs with ca. 11 organs ( Fig. 109 View FIGURES 105 – 109 ). Articles of ultimate legs inflated. Ratio of length of telopodites of ultimate legs/length of metasternite ca. 4.23: 1. Shape and chaetotaxy as in Figs. 108, 109 View FIGURES 105 – 109 .
Postpedal segments. Intermediate tergite with posterior margin convex ( Fig. 108 View FIGURES 105 – 109 ); intermediate sternite with posterior margin slightly concave, ( Fig. 109 View FIGURES 105 – 109 ); posterior border of first genital sternite very slightly convex ( Fig. 109 View FIGURES 105 – 109 ). Gonopods uniarticulate, not separated on the middle ( Fig. 109 View FIGURES 105 – 109 ). Anal organs absent.
Variation. Females with 47, 49, (probably 51), 53 leg-bearing segments; males with 47, 49, 51.
All other characters without significant variation.
Remarks. An accurate comparison of the characters of specific value in the lectotype male ( Figs. 99–104 View FIGURES 99 – 104 ) and paralectotype female ( Figs. 105–107 View FIGURES 105 – 109 ), with those of the specimens cited in "Other material examined" ( Figs. 54– 98 View FIGURES 54 – 58 View FIGURES 59 – 66 View FIGURES 67 – 74 View FIGURES 75 – 80 View FIGURES 81 – 88 View FIGURES 89 – 94 View FIGURES 95 – 98 , 108, 109 View FIGURES 105 – 109 ) leave no doubts as to the conspecificity of all of them. The precedent redescription of P. zapallar is based on the non-type specimens, because the lectotype male is incomplete and the paralectotype female shows a sub-adult condition, indicated by the relatively small body size, empty spermathecae (located at level of the penultimate leg-bearing segment), low number of organs in each cluster of coxal organs and reduced pilosity on the articles of ultimate legs.
Plateurytion zapallar was inadequately described by Chamberlin; his original description does not refer to a specimen in particular, nor does it specify the sex of the studied specimens. It only includes a single inaccurate figure of the cephalic plate, and completely lacks information on pilosity of the antennae; kind and number of specialized sensilla of a.a. II, V, IX and XIII; shape of mandibles; anterior and posterior limits of ventral porefields; shape of postpedal segments; etc.
In his description Chamberlin states "First joint of telopodite of first maxillae bearing a sensory lappet, the syncoxite with none", but the coxosternite of the first maxillae actually bears lappets ( Figs. 99–101 View FIGURES 99 – 104 : a). About the ventral pores, the author says "Ventral pores present in a circular area in front of caudal margin of sternite", this is correct for the metasternites of the anterior region of the body, but in those of the posterior region, the pore-fields are divided in two subsymmetrical areas.
Type locality. CHILE: Aconcagua: Zapallar . (According to the current Chilean territorial organization, Zapallar belongs to Petorca province).
Known range. CHILE: Region IV (Coquimbo region): Elqui province: 11 Km N of Los Vilos. Region V (Valparaíso region). Petorca province: Zapallar ; Quebrada Huaquén, Pichicuy. Quilota province: La Campana National Park. Valparaíso province: Quebrada el Tigre, Cachagua. (See Fig. 110 View FIGURE 110 ).
Remarks. According to the biogeographical regionalization of the Andean Region proposed by Morrone (2015), the geographical distribution of this species corresponds to the "Coquimban province" and "Santiagan province" (Central Chilean sub-region).




| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chilerium zapallar
| Pereira, Luis Alberto 2015 |
zapallar
| Pereira 2008: 56 |
| Bonato 2007: 6 |
Eurytion zapallar:
| Pereira 2006: 167 |
| Foddai 2000: 75 |
Chilerium zapallar
| Chamberlin 1955: 24 |