
Diomus lupusapudoves, Vandenberg & Iverson & Liere, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4420.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:F3B2F64C-5896-4896-846A-E3B2F8B338FF |
|
DOI |
https://doi.org/10.5281/zenodo.5968649 |
|
persistent identifier |
https://treatment.plazi.org/id/886E87D0-FFD7-EB13-F1C9-C5A2FBC53646 |
|
treatment provided by |
Plazi |
|
scientific name |
Diomus lupusapudoves |
| status |
sp. nov. |
Diomus lupusapudoves , sp. nov.
( Figs. 1–7 View FIGURES 1–3 View FIGURES 4–8 , 9 View FIGURES 9–10 , 11–15 View FIGURES 11–13 View FIGURES 14–15 )
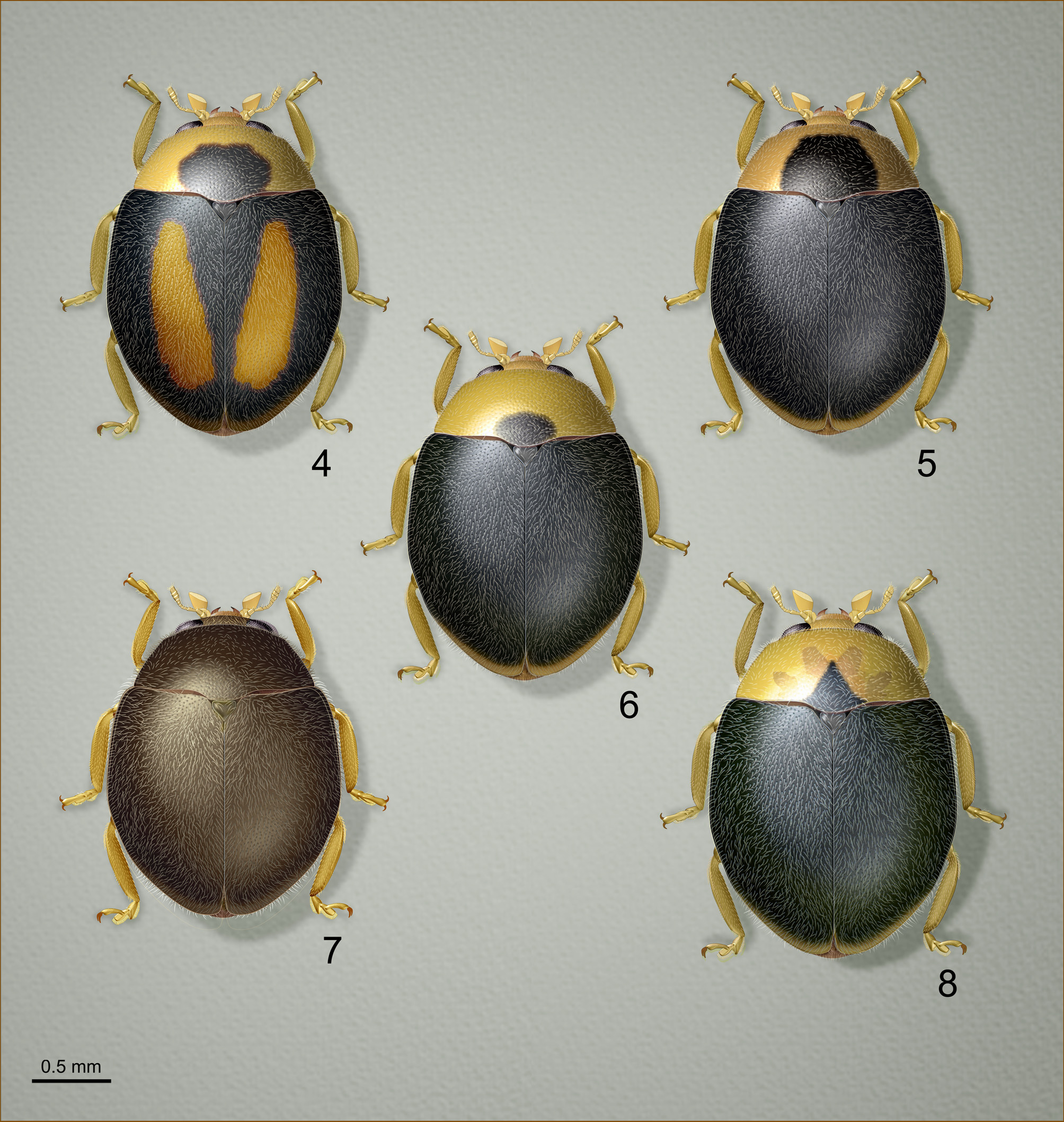
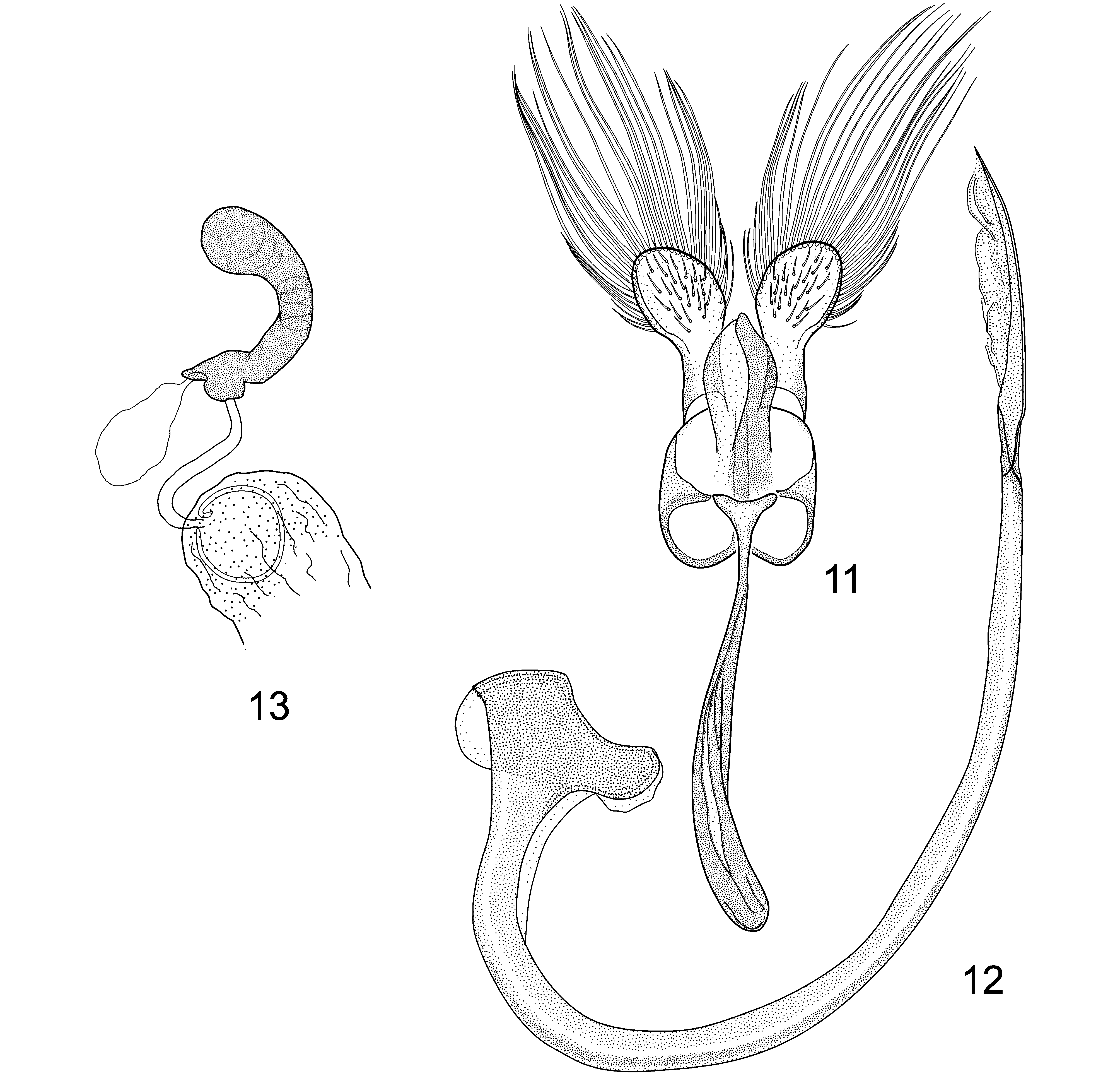
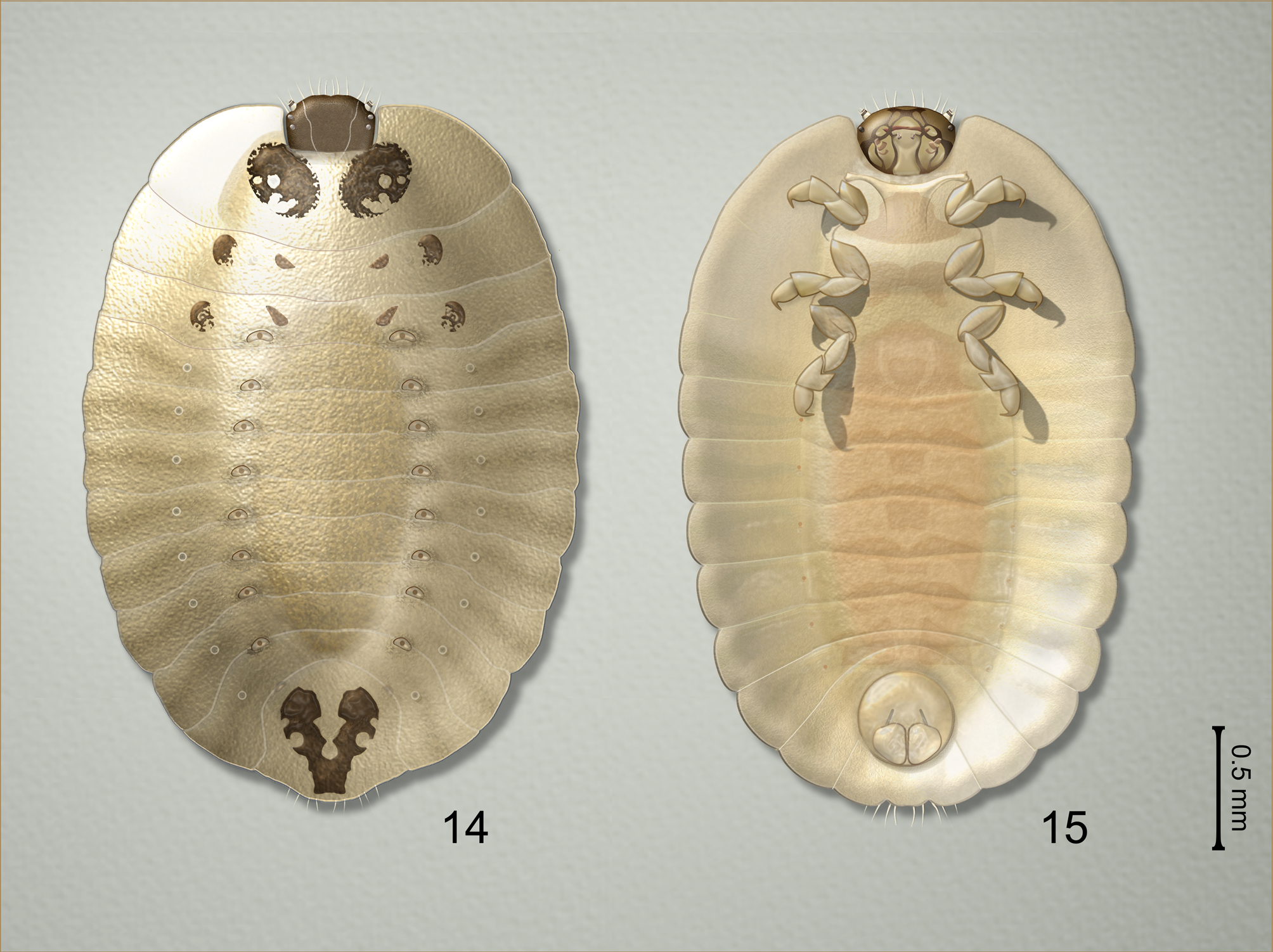
Diagnosis. Due to variability in the dorsal color pattern ( Figs. 4–7 View FIGURES 4–8 ), the new species is best identified by the exact configuration of the adult male genital structures ( Figs. 11–12 View FIGURES 11–13 ) and by the myrmecophilous, onisciform (=platyform) larva ( Figs. 14–15 View FIGURES 14–15 ) possessing a finely granulate dorsum devoid of setae, and with the epipleurum of each abdominal segment extended into an oblique lateral plate, forming a protective skirt-like border to shield the underside of the body and appendages.
This species closely resembles another myrmecophile, Diomus thoracicus (Fabricius, 1801) , recorded from northern South America and the Antilles, to Mexico and southern Florida ( Peck 2015), but the adult of our new species is smaller on average (1.7–2.1 mm vs. 2.0– 2.4 mm). Diomus thoracicus male genitalia (illustrated in Gordon 1999) are structurally quite similar as well, but possess at least 1.5× the number of setae along the outer margins of each paramere and have the trabes strongly inflated in apical 2/3. The larva of D. thoracicus is also onisciform ( Vantaux et al. 2010; Roux et al. 2017), but has the dorsal surface a uniform light bluish gray except for the contrasting white epipleurum of each abdominal segment. The perimeter of the body is distinctly fimbriate in D. thoracicus , whereas the new species has only a few setae on the head and posterior end of the abdomen. In addition, the head of the D. thoracicus larva projects anteriorly, and is not enclosed laterally by the sides of the pronotum as it is in the new species. Although larvae of both species are myrmecophilous, D. thoracicus larvae are intranidal parasites that feed on ant brood, whereas larvae of the new species are coccidophagous and occur on open vegetation.
Another species, D. urban Gordon, 1999 ( Fig. 8 View FIGURES 4–8 ), was found infrequently alongside the new species among colonies of C. viridis in the coffee agroecosystem that we studied in Chiapas, Mexico. Males and females of D. urban are superficially very similar to the unspotted male form of the new species ( Figs. 5–6 View FIGURES 4–8 ), but actually belong to an entirely different group within Diomus ( Diomus Group G, sensu Gordon 1999) characterized by the male genitalia with extremely wide and densely setose, funnel-shaped parameres, and the penis with an apical flagellum. Females belonging to this group are distinguished by a heavily sclerotized, cylindrical or spindle-shaped nodulus and elongate tapered cornu of the spermathecal capsule. Externally, adults of D. urban can be separated from the new species by their slightly more robust form and the longer intercoxal process of the prosternum ( Fig. 10 View FIGURES 9–10 ) with the carinae more narrowly separated posteriorly and more weakly convergent anteriorly.
Description of holotype (adult, male) ( Fig. 4 View FIGURES 4–8 ). Length 2.0 mm, width 1.5 mm. Form broadly oval, convex (height = 0.9 mm). Metathoracic wing present. Dorsal surfaces finely granulate, feebly shining, distinctly punctate, pubescent. Coloration of head including mouthparts pale yellow with mandibular apex reddish amber; pronotum pale yellow with large irregularly rounded basomedian reddish brown spot. Elytron predominantly black with faint bluish sheen, with diffusely lighter yellow brown elytral apex; disc bearing large teardrop shaped oblique reddish orange mark; elytral epipleuron yellow brown with margins narrowly dark brown. Scutellar shield dark brown. Venter predominantly brown, darkest on meso-, metaventrite; prosternum pale yellow with median half yellow brown; mesepimeron paler than surrounding sclerites with margins narrowly darkened; abdomen pale brown, lighter, yellower toward apex. Legs pale yellow with meso-, metacoxae slightly darker. Pubescence shiny off white.
Dorsal punctation moderately coarse; punctures separated by approximately 1× diameter on elytron, equal in width to eye facet; punctures slightly coarser, more crowded on head, pronotum. Dorsal pubescence semierect, moderately dense, evenly distributed, lacking distinct setal pattern; individual setae equal to about 1/3 to 3/4× length of scutellar shield, arcuate; elytral setae directed more or less posteriorly except directed outwardly at sides of body, posteromedially near suture.
Head large, 0.64× width of pronotum; eye large, finely facetted, with interfacetal setae, distinctly notched near antennal insertion by small triangular ocular canthus; inner orbits evenly arcuate, with minimum separation near midlength; interocular distance 1.6× width of eye in frontal view. Antenna composed of 11 antennomeres; antennomere 3 elongate, about 2.3× length of antennomere 4; last four antennomeres subequal in length, forming gradual oval club expanding apically from antennomere 8–10; antennomere 11 subrhomboidal, slightly narrower, than 10. Maxillary palp with terminal palpomere strongly expanded distally, triangulate, about as long as greatest width. Pronotum with basal width about 2× length, tapered toward apex, strongly convex. Elytron in dorsal view with lateral margin evenly arcuate; elytral apices dehiscent. Ventral surfaces pubescent, distinctly punctate; punctation less even, generally less dense than elytral punctation. Prosternum ( Fig. 9 View FIGURES 9–10 ) with intercoxal process short, broad, with convergent carinae extending to, merging near apex, framing ovotriangular depression. Suture between abdominal ventrites 1 and 2 distinct, linear; 5th ventrite with apex shallowly, roundly emarginate; 6th ventrite with apex bearing indistinct shallow notch at middle. Tarsal claw with broad scythe-like inner tooth extending from base to apical 1/3 or beyond.
Male genitalia ( Figs 11–12 View FIGURES 11–13 ). Basal lobe (=penis guide sensu Ślipiński 2007) moderately flat, in ventral view expanding from base to apical 2/5, tapered beyond to pointed apex, asymmetrical, about 2/3 length of paramere. Paramere with apical 1/2 roundly expanded, with about 19–25 long setae along perimeter, with few shorter setae in more proximal position on each side; setae slightly flattened, broad at base, sharply pointed distally; inner (=ventral) surface of paramere with very short setae sparsely distributed. Penis (=aedeagus or sipho) with outer arm of capsule short, truncate, bearing semicircular crest on outer surface; inner arm short, roundly tapered; penis apex lacking flagellum, with slightly expanded membranous area.
Female ( Fig. 7 View FIGURES 4–8 ). Similar to male except body form proportionally shorter, dorsal coloration uniformly medium brown. Prosternum and underside of head yellow brown. 5th abdominal ventrite with posterior margin roughly linear in median 1/3; 6th ventrite with posterior margin arcuate. Tarsal claw with short triangular tooth near base, not extending beyond apical 1/2. Spermathecal capsule of female genitalia ( Fig. 13 View FIGURES 11–13 ) moderately small, weakly bent near middle; nodulus (=collum) in the form of short annular projection; ramus sessile, with thorn-like apodeme (=beak sensu Gordon 1999) above accessory gland; cornu short, distinctly swollen distally. Sperm duct lacking sclerotized sheath or process (=infundibulum auctorum). Bursa with lightly sclerotized internal chamber (=bursal plate sensu Gordon 1999) visible as light brown ring-like structure.
Variation ( Figs 4–7 View FIGURES 4–8 ). Length 1.7–2.1 mm. Male with basal mark on pronotum variable in size; reddish orange oblique mark on elytron may be absent or only faintly indicted as slightly lighter area on disc; pale band at elytral apex yellow to brown or indistinct. Female with dorsal coloration light brown to blackish, unicolorous or with anterior pronotal angles and extreme elytral apex defusely lighter.
Larva (based on field collected 2nd–4th instars) ( Fig. 2 View FIGURES 1–3 , 14-15 View FIGURES 14–15 ). Form broadly oval, onisciform. Body light grayish tan with head, thoracic and terminal abdominal plates medium brown. Anterior margin of head, posterior margin of abdominal apex with few simple setae. Dorsum devoid of setae, with granulate texture consisting of tiny fingerprint-like ridges and pits (visible at 50× magnification or greater), with eight conspicuous pairs of intersegmental pores near anterior borders of abdominal segments 1–8; first pair situated more anteriorly, appearing to extend onto base of metanotum. Head with frontal arms of epicranial suture well developed; epicranial stem obsolete. Pronotum with pair of pigmented plates; external pronotal margins expanded, flattened, surrounding sides of head. Meso-, metanotum similarly expanded, each with two pairs of small pigmented plates. Abdominal segments transverse, with expanded, flattened epipleurum; tergum 9 with v-shaped pigmented plate. Venter concave, without strongly sclerotized plates; legs short, robust, not capable of extending beyond sides of body; pygopod well developed.
Pupa ( Fig. 3 View FIGURES 1–3 ). Form broadly oval, attached at caudal end, not enclosed in shed larval skin; surface bearing glandular hairs. Coloration yellow to yellowish brown.
Trophic relations. Larvae and adults have been observed feeding on Coccus viridis (Green) on coffee plants, especially those tended by Azteca sericeasur Longino and Pheidole synanthropica Longino ants.
Etymology. The species name is a concatenation of the Latin phrase “lupus apud oves” (=wolf among the sheep), in reference to its presence among coccids tended by ants.
Type material. Holotype (male) with labels: “MEX: Chiapas: Finca Irlanda, 15.173583°−92.336081°, 28.July.2010, coll. H. Liere / On Coffea arabica plants among C. viridis , newly eclosed adult / HOLOTYPE, Diomus lupusapudoves Vandenberg, Iverson & Liere, 2018 ” (USNM); 19 paratypes (9 females, 10 males): 2 with same labels as type except third label “PARATYPE, Diomus lupusapudoves Vandenberg, Iverson & Liere, 2018 ” (USNM); 3 with labels “MEX: Chiapas: Finca Irlanda, 15.173583° −92.336081°, 24.May.2010, coll. A. Iverson / On Coffea arabica plants / PARATYPE, Diomus lupusapudoves Vandenberg, Iverson & Liere, 2018 ” (USNM); 14 with labels “MEX: Chiapas: Finca Irlanda, 15.173583° −92.336081°, 10.June.2010, coll. A. Iverson / Reared from late instar onisciform larva collected on Coffea arabica plants / PARATYPE, Diomus lupusapudoves Vandenberg, Iverson & Liere, 2018 ” (13 USNM, 6 ECO-TAP-E)
Other material examined. A small amount of additional material of the new species bearing the same collection data as the 24.May.2010 and 10.June.2010 paratypes has been deposited in the USNM. These represent less well preserved exemplars (either disarticulated, fragmented, or rubbed), but still deemed useful for studying certain morphological or anatomical details. Specimens from each collection event have been placed together on a single pin using multiple points and/or within a gelatin capsule. A vial of mixed second through fourth instar larvae of the new species from the 10.June.2010 collection event has been deposited in the USNM alcohol collection. A male and a female specimen of D. urban from the 24.May.2010 collection event have been point-mounted and deposited in the USNM dry collection. The later collection also contains a large quantity of material of other South American Diomini documented in the Gordon (1999) monograph of this group and utilized for comparative purposes during our study.
Remarks. Gordon (1999) informally divided the genus Diomus into eight species groups ( Diomus groups A through H) based almost entirely on male genitalic characteristics. He indicated that some of these groups may be paraphyletic, but still have a value for identification purposes. Using his system, our new species belongs in group B and appears to be closely allied to D. thoracicus , the type species of the genus.
Our new species exhibits strong sexual dimorphism with respect to the dorsal color pattern ( Figs. 4–7 View FIGURES 4–8 ). This situation is not uncommon in the tribe Diomini , and probably explains why the females of many other species have not been associated with their male counterparts. We have managed to include females in our type series because both sexes were reared together from the same distinctive onisciform larvae, and have the same configuration of the antenna, prosternal process, punctation, and pubescence (dorsal setal pattern).


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.