
Bonnetina juxtantricola, Ortiz & Francke, 2014
|
publication ID |
https://doi.org/ 10.1080/00222933.2014.924770 |
|
DOI |
https://doi.org/10.5281/zenodo.4327863 |
|
persistent identifier |
https://treatment.plazi.org/id/891E87F1-FF94-FF85-FE38-806CFBB0FDCC |
|
treatment provided by |
Carolina |
|
scientific name |
Bonnetina juxtantricola |
| status |
sp. nov. |
Bonnetina juxtantricola View in CoL sp. nov.
urn:lsid:zoobank.org:act:7A6A6CC2-E07C-4CFD-910D-0255D69F7AD4
( Figures 9–13 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 ; Table 1)
Holotype. ♂ ( CNAN-T0758 ). MEXICO: Guerrero state: Quechultenango municipality: Outside of Juxtlahuaca Cavern : 17.4387°, −99.1595°: 930 m asl. 12 September 2005. Alejandro Valdez & Héctor Montaño, cols. Under a stone.
Paratype. ♀ ( CNAN-T0759 ): same locality, collectors and microhabitat as holotype. 19 July 2005 .
Etymology
The specific name is composed from the Latin preposition juxta (nearby), the noun antrum (cavern) and the suffix cola (dweller). The name can be translated as ‘nearbycavern dweller’ and makes reference to the type locality of this species, which is the surroundings of an archaeologically very valuable cave.
Diagnosis
Males differ from those of all known Bonnetina species in the shape of the palpal bulbs. Additionally, from B. rudloffi , B. alagoni , B. papalutlensis , B. tenuiverpis and B. tanzeri in that PI bulbal keel is not denticulate as on those species. It also differs from males of B. aviae in the lesser retrolateral bend of the embolus, which describes an angle of approximately 30° with the axis of the bulb and about 70° in B. aviae . The male tibia I accessory apophysis is present but considerably less developed than that of B. cyaneifemur , B. rudloffi , B. papalutlensis , B. tanzeri and B. tenuiverpis . Females have domiform spermatheca and so differ from the species that have it digitiform ( B. cyaneifemur ), subdigitiform ( B. tenuiverpis ), subtriangular ( B. papalutlensis ) or flattened ( B. tanzeri ). Females differ from those of B. aviae and B. alagoni by presenting very distinctly thick, erect setae on the posterior area of carapace.
Description
Male holotype
Morphology
Some quantitative characters are given in Table 1
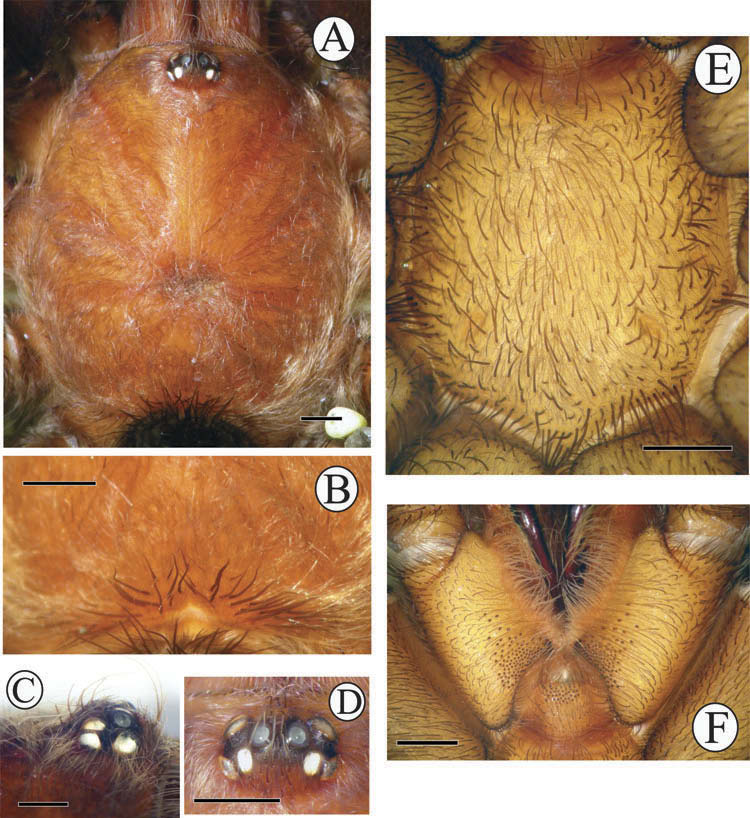
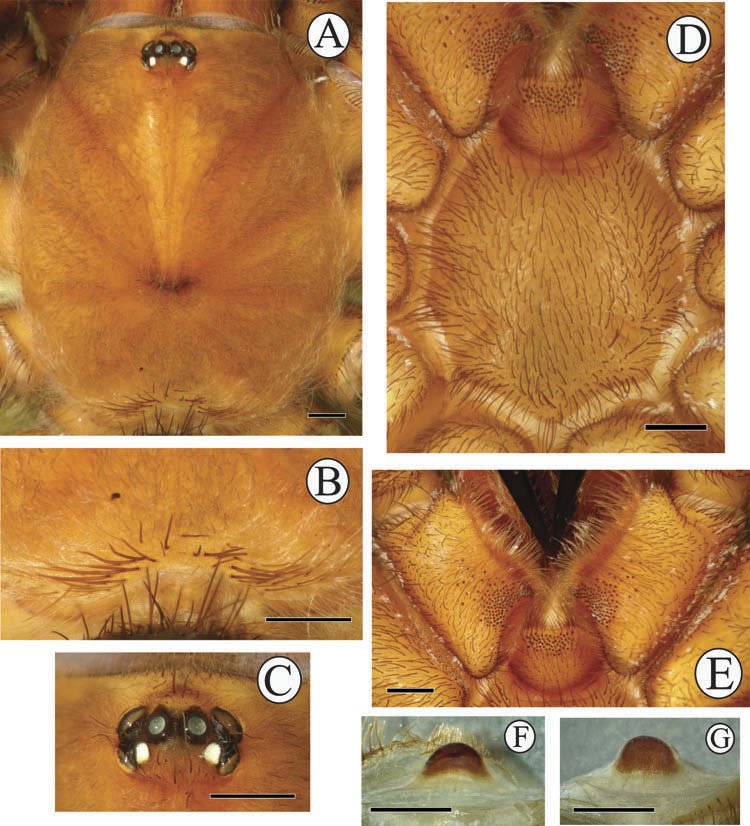
Carapace with caput nearly flat; fovea broad and procurved ( Figure 9A View Figure 9 ). Posterior area of carapace bears distinctly thick erect setae ( Figure 9B View Figure 9 ). Ocular area: eight eyes disposed in two rows on elevated tubercle (PLE oblique to carapace) ( Figure 9C View Figure 9 ); anterior eye row procurved; posterior row recurved ( Figure 9D View Figure 9 ). Ocular quadrangle width, 1.36; length, 0.94. Clypeus, 0.32 wide. AME circular, diameter, 0.36; ALE ovoid, greater diameter, 0.40; PME ovoid, greater diameter, 0.34; PLE ovoid, greater diameter, 0.34.
Chelicerae with seven teeth (left appendage) and seven teeth plus one tiny apicalmost tooth (right), close and parallel to the promargin of ventral side.
Sternum ( Figure 9E View Figure 9 ) slightly convex to its centre, covered uniformly by thick erect hairs and other hairs much smaller; with three pairs of sigillae, placed opposite to coxae I, II and III. Labium subtrapezoidal; middle length, 0.96; anterior width, 0.90; posterior width, 1.70 ( Figure 9F View Figure 9 ).
Appendage segment lengths (left limbs). Palp: femur, 4.9; patella, 3.2; tibia, 4.6; total,12.7. Leg I: femur, 8.1; patella, 4.3; tibia, 6.0; metatarsus, 5.6; tarsus, 4.2; total, 28.2. Leg II: femur, 6.7; patella, 4.1; tibia, 5.4; metatarsus, 5.2; tarsus, 3.8; total, 25.2. Leg III: femur, 6.1; patella, 3.3; tibia, 4.5; metatarsus, 6.3; tarsus, 3.9; total, 24.1. Leg IV: femur, 8.2; patella, 3.8; tibia, 6.6; metatarsus, 9.1; tarsus, 4.5; total, 32.2 (Leg formula: leg IV> I> II> III).
Retrolateral face of palpal tibiae with prominent, apically inclined, cone-shaped nodule near apex, covered by long thick setae.
Palpal bulbs ( Figures 10 View Figure 10 , 11 View Figure 11 ) robust with thick embolus, that is moderately curved and twisted dorsally and retrolaterally (from base to apex) so that in the apex, the ventral structures of the bulb become prolateral. Prolateral inferior, prolateral superior, sperm pore and prolateral sub-apical keels present. PI keel is smooth and extends from close to the base of the embolus to near to sperm pore; it ends sharply at its apex, forming a distinct prominence. PS keel extends from embolus base to apex. SP keels extend from the bulb apex to the sperm pore and are folded onto each other, except in the basalmost region; they are fused at the apex, not forming a pore at the embolus tip. PSA keel, about as long as SP keels, is subapical to the embolus and located between PS and sperm pore.
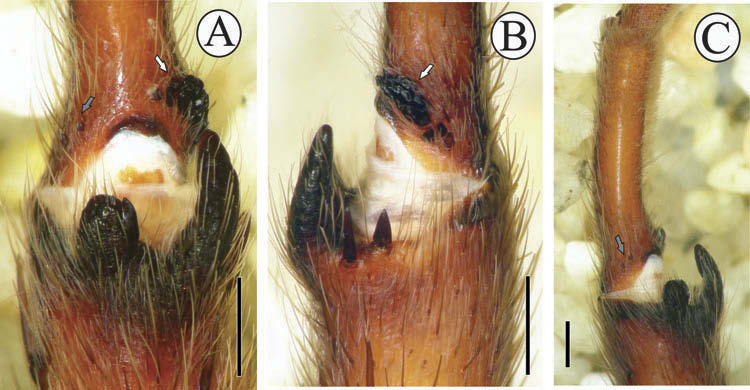
Legs I holding organ. Tibiae I with three apophyses near the apex, all parallel to segment axis ( Figure 12A, B View Figure 12 ). Prolateral apophysis tubular, blunt at its apex and bearing a megaspine on its internal border; length, 0.48/0.48 (left/right). Retrolateral apophysis curved, acute at apex; length, 1.20/1.10. Accessory apophysis very poorly developed, bearing a subtriangular megaspine at its apex and a spine on the internal border; length, 0.14/0.10. When flexed, the moderately curved metatarsus I ( Figure 12C View Figure 12 ) folds between prolateral and retrolateral apophyses. Metatarsi I with nodule of 16 (left) and 19 (right) granules on basal ventro-retrolateral region ( Figure 12B View Figure 12 ). Two (left) and one (right) small rounded structures, presumably modified spines, on the basal ventro-prolateral region of metatarsi I ( Figure 12A, C View Figure 12 ).
Femora of palps and legs I and II prolaterally and femora of legs IV retrolaterally covered by pads of ciliated hairs.
Palpal coxae and trochanters with non-plumose setae pro- and retrolaterally.
Scopulae Metatarsi. On legs I entire segment except the basal 1/5; on legs II apical 2/3; on legs III on apical half (left) and apical 1/3 (right) of segment; on legs IV apical 1/5. Tarsi. On legs I and II entire with few dispersed type B hairs; on legs III divided completely by a band of 1–3 fine hairs; on legs IV divided full length by band of 2–4 very thick hairs.
Claw tufts dense on every leg.
Abdominal urticating hairs. Type III, in patch located dorsally in central-posterior half of abdomen, covering 0.31 of its length. It could have been oval or subrectangular, according to the mark, but only few hairs remain, located on its anterior border.
Spination pattern (left limbs). Palp: femur p0-0-1; tibia p1-2-0. Leg I: femur p0-0-1; tibia v3-2-2 p0-1-1; metatarsus v0-0-1 p0-0-1. Leg II: femur p0-0-1; tibia v3-2-3 p0-1-1 r1-0-0; metatarsus v2-1-3 p0-1-1. Leg III: femur p0-1-1; tibia v1-2-2 p0-2-1 r1-1-1; metatarsus v2-3-3 p1-1-2 r0-1-1. Leg IV: femur r0-0-1; tibia v1-2-3 p0-1-1 r0-2-1; metatarsus v10 p1-2-1 r0-1-1.
Preservation state
The specimen is in good condition, stored in a flask of 80% ethanol. Both pedipalpal bulbs are housed apart, coated with gold. The specimen is homogeneously reddishbrown due to the length of time it has remain preserved.
Allotype female
Morphology
Some quantitative characters are given in Table 1
Carapace ( Figure 13A View Figure 13 ) with caput elevated. Fovea procurved. Posterior area of carapace bears distinctly thick erect setae ( Figure 13B View Figure 13 ). Anterior eye row clearly procurved; posterior row recurved ( Figure 13C View Figure 13 ). Ocular quadrangle width, 1.44; length, 0.94. Clypeus, 0.22 wide. AME circular, diameter, 0.36; ALE ovoid, greater diameter, 0.52; PME ovoid, greater diameter, 0.30; PLE ovoid, greater diameter, 0.34.
Chelicerae with nine teeth plus one tiny basalmost tooth (left appendage) and eight teeth plus one tiny basalmost tooth (right), close and parallel to the promargin on ventral side.
Sternum ( Figure 13D View Figure 13 ) slightly convex, covered uniformly by erect thick hairs and other hairs much smaller; with three pairs of sigillae, placed opposite to coxae I, II and III. Labium ( Figure 13E View Figure 13 ) subtrapezoidal; middle length, 1.30; anterior width, 0.96; posterior width, 2.02.
Appendage segment lengths (left limbs). Palp: femur, 5.7; patella, 3.5; tibia, 3.9; tarsus, 4.1; total, 17.2. Leg I: femur, 7.8; patella, 5.0; tibia, 5.3; metatarsus, 4.1; tarsus, 2.3; total, 24.5. Leg II: femur, 6.6; patella, 3.7; tibia, 4.7; metatarsus, 4.0; tarsus, 3.1; total, 22.1. Leg III: femur, 6.0; patella, 3.9; tibia, 4.1; metatarsus, 5.3; tarsus, 3.3; total, 22.6. Leg IV: femur, 8.0; patella, 4.3; tibia, 6.0; metatarsus, 7.8; tarsus, 3.7; total, 29.8 (Leg formula: leg IV> I> III> II).
Only one spermatheca, domiform in shape ( Figure 13F, G View Figure 13 ). It is fully sclerotized ventrally, while dorsally apically only, forming a crescent-shaped figure. Length, 0.46; base width, 0.80.
Scopulae Metatarsi. Legs I: entire on proventral half, 3/4 apical on retroventral half. Legs II: 3/4 apical on anterior half, 1/4 apical on posterior half. Legs III: 1/3 apical. Legs IV: 1/5 apical. Tarsi. As in holotype .
Palpal coxae and trochanters with non-plumose setae pro- and retrolaterally.
Femoral pads as in holotype.
Abdominal urticating hairs. Type III, in oval dorsal patch (with rear part moderately narrower), located dorsally on central-posterior half of abdomen, covering 0.33 of its length .
Spination pattern (left limbs). Palp: femur, p0-0-1; tibia, v0-3-3 p0-1-0. Leg I: femur, p0-0-1; tibia, v0-1-0; metatarsus, v0-1-1 p0-0-1. Leg II: femur, p0-0-1; tibia, v0-1-4 p0- 1-1; metatarsus, v1-1-2 p0-1-0. Leg III: femur, p0-0-1; tibia, v2-2-4 p1-1-0 r1-1-1; metatarsus, v0-3-5 p1-2-2 r0-1-1. Leg IV: femur, r0-0-1; tibia, v1-2-3 p0-1-1 r1-0-2; metatarsus, v9 p0-2-1 r0-2-1.
Preservation state
The specimen is in good condition, stored in a flask of 80% ethanol. Genital area is housed in a plastic vial inside the flask. The specimen is homogeneously reddishbrown, due to the length of time it has remain preserved.
Distribution
Bonnetina juxtantricola is known only from the vicinity of Juxtlahuaca cavern, situated in Guerrero state and geographically at 930 m asl in the Sierra Madre del Sur, a mountain range that extends parallel to the Pacific Ocean coast between Central and Southern Mexico.
Natural history
Very little is known of the natural history of this species. The only known male was collected in September, suggesting that this month could be part of its breeding season. Both male and female were collected under stones in a tropical deciduous forest.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theraphosinae |
|
Genus |