
Heligmonoides mirzai, Smales, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.5341456 |
|
persistent identifier |
https://treatment.plazi.org/id/8B20878D-FFD0-FFCE-E2BE-FA0FFD6FFCFF |
|
treatment provided by |
Diego |
|
scientific name |
Heligmonoides mirzai |
| status |
sp. nov. |
Heligmonoides mirzai , new species
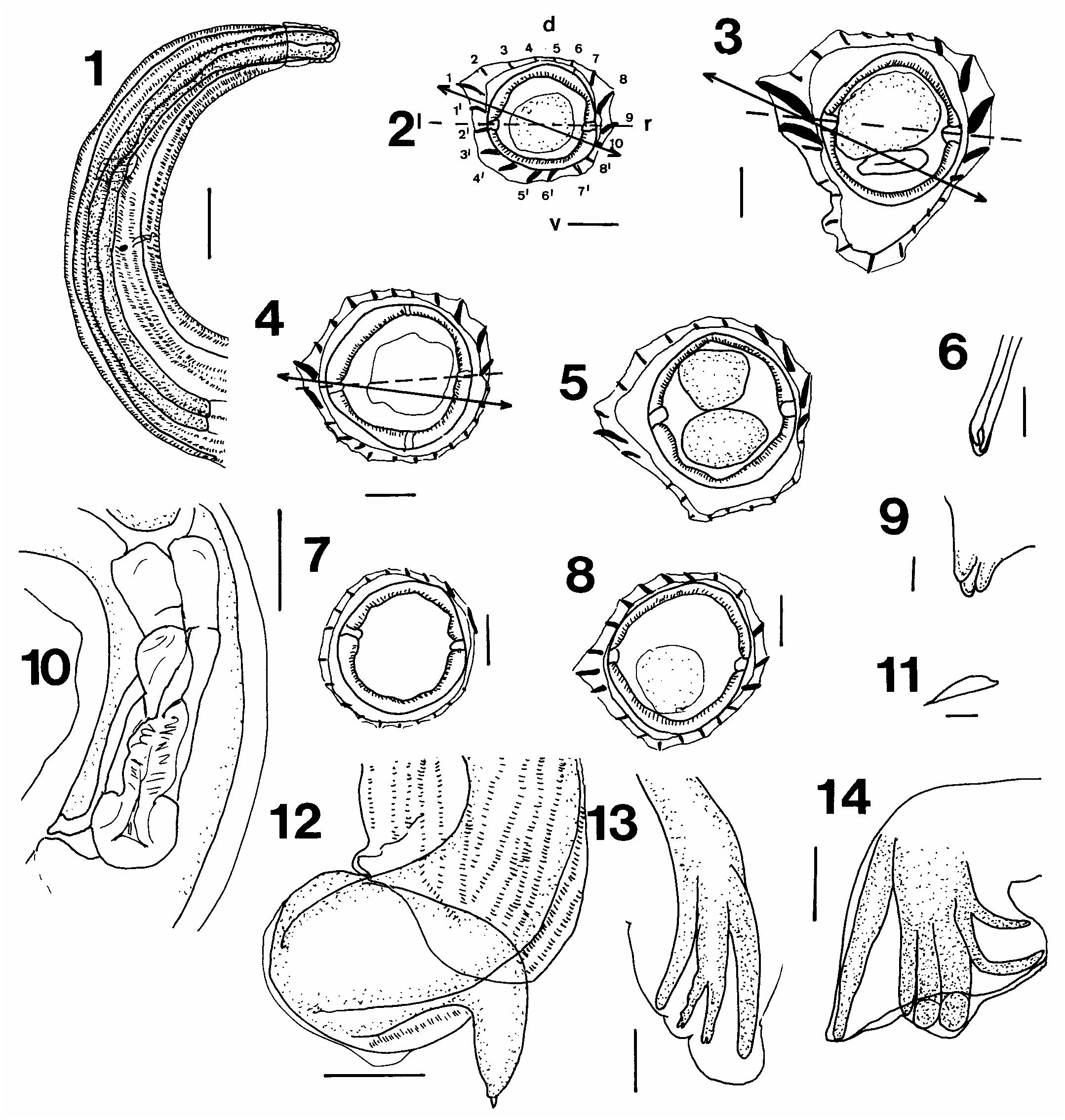
( Figs. 1–14 View Figs )
Material examined. – Holotype male and allotype female: BBM- NG-99529, from the small intestine of Melomys rufescens ( Muridae : Hydromyinae ), type host, Nauti Village, 7°15'S 146°13'E, Morobe Province, Papua New Guinea, coll. A. B. Mirza, 10 Oct.1970.
Paratypes: 12 males, 12 females, SAM AHC 34783; collection data as above .
Voucher specimens: BBM NG- 97928 1 male, 2 females from the small intestine of Melomys rufescens, Minava Village, Watut , 6°50'S 146°21'E, Morobe Province, coll. A. B. Mirza, 23 Oct.1969 GoogleMaps ; AM W32620, 2 males, 3 females from small intestine of Melomys rufescens, Yapsiei area, 4°35'S 141°5'E, Sanduan Province, coll. T. Flannery, 17 Jan.1984 GoogleMaps ; 1 male, 2 females, AM W32625 from the small intestine of Melomys species , Dokfuma Star Mountains 5º01'S 141°07'E, West Sepik, Sanduan Province, coll. T. Flannery, 5 Apr.1984 GoogleMaps .
Prevalence. – A total of 5 of 25 M. rufescens and 1 of 10 Melomys spp.
Etymology. – The species name is in recognition of the extensive collecting done by A. B. Mirza in the 1960s and 1970s in New Guinea.
Description. – Small coiled worms; prominent cephalic vesicle present; buccal capsule vestigial. Mouth opening triangular with rudimentary lips; labial and cephalic papillae not observed. Oesophagus claviform. Nerve ring surrounds oesophagus at about middle third; deirids and excretory pore at about same level, posterior to nerve ring. Synlophe of pointed longitudinal cuticular ridges extends from the posterior margin of the cephalic vesicle to level of spicules or vulva; 16–18 ridges in anterior, 21–24 in midbody. Axis of orientation of ridges passing from ventral right to dorsal left side, inclined about 75° from sagittal axis; 10–12 ridges dorsal side, 11–12 ridges ventral side in midbody. Ridges 1, 1’ large but not thickened basally, together with ridges 2, 2’ form a carene of type B. Ridges 3–7 decreasing in size, ridges 8–10 increasing in size, ridge 11 smaller; ridges 3’– 8’ increasing in size, ridges 9’–11’ decreasing in size. Posterior region of body with 19–20 (male) 20–21 (female) ridges reduced in size; 10–11 dorsal side, 9–10 ventral side ridges.
Male (holotype and 9 paratypes): Length 2.2–3.3 (2.8) mm, maximum width 83–107 (90). Cephalic vesicle 36–46 (39.25) long. Oesophagus 350–550 (420) long; nerve ring 145 from cephalic end. Bursa asymmetrical, left lobe larger, rays of left lobe more robust; without dorsal median notch. Dorsal ray symmetrical; divided at about half its length, each branch dividing again at distal tip; terminal divisions, rays 9, 10 symmetrical; rays 8 arising at same level proximally to division of dorsal ray, left ray 8 slightly more robust than right. Rays 4, 5, 6 with common stem, reaching margin of bursa, rays 6 more slender than rays 4, 5. Rays 2, 3 with common stem, widely divergent distally, rays 3 longer than rays 2. Genital cone short, lightly sclerotized, ventral lobe with unpaired papillae 0, dorsal lip bifid, each lobe with single papilla 7. Spicules equal, filiform, proximal ends rounded, distal tips simple, straight, 250–320 (275) long. Gubernaculum 27 long.
Female (allotype and 9 paratypes): Length 2.95–3.7 (3.4) mm, maximum width 99–107 (104). Cephalic vesicle 42.0 –49.5 (45) long. Oesophagus 350–400 (380) long; nerve ring 190, 220, excretory pore 250, 280 deirids 250, 280 from cephalic end. Posterior end twisted and flexed ventrally, with praepuce in mature worms; vulva opening about 100 from tail tip. Monodelphic; ovejector with sphincter, 95, longer than vestibule 40, and infundibulum, 45. Tail conical, tip with spike 39.5–55 (48.5) long. Eggs thin- shelled, ellipsoidal, in utero 51–69 (57) by 30–36 (33.8).
Remarks. – Despite the fact that much of the material had been fixed in the hosts in situ, making it difficult to discern the morphology of the anterior end of the worms, it could be seen that Heligmonoides mirzai , new species, has most of the morphological features of the genus Heligmonoides and resembles H. bulbosus Ow Yang et al., 1983 , in the number and arrangement of the ridges on the synlophe ( Ow Yang et al., 1983, Fig 11A, B View Figs ). Heligmonoides mirzai , new species, difers from H. bulbosus in spicule length (250–320 compared with 92–100), the proportions of the dorsal ray, the length of rays 3 and the shape of the female posterior end, with a praepuce and spiked tail tip.
Hasegawa & Syafruddin (1994a) originally described Maxomystrongylus musseri as Heligmonoides musseri , noting that it most closely resembled H. bulbosus , particularly in the features of the carene; supported by 3–4 slender ridges lacking thickened roots. Subsequently these authors erected the new genus Maxomystrongylus Hasegawa & Syafruddin, 1997 , and transferred H. musseri into it on the basis of the characters of the synlophe discussed above, having the division of the dorsal ray distal to the derivation of rays 8 and a vagina vera with a unilateral diverticulum (Hasegawa & Syafruddin, 1994a). Further they commented that the systematic relationship between H. bulbosus and Maxomystrongylus remained unclear. Although H. mirzai , new species, has characters of the synlophe and a form of the dorsal ray consistent with Maxomystrongylus , it does not have a unilateral diverticulum of the vagina. Consequently it has been placed in the genus Heligmonoides together with the other Southeast Asian representatives of the genus, H. bulbosus and H. lanceolatus Ow Yang et al., 1983 , pro tem. A revision of the heligmonellid genera from the Oriental and Australasian regions is needed to resolve the relationship between Heligmonoides and Maxomystrongylus and the systematic position of the Australasian species.
| SAM |
South African Museum |
| AM |
Australian Museum |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |