
Canthyporus reebae, Manuel, Michaël & Ramahandrison, Andriamirado T., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4273.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:F42A8DE1-C70E-47E1-AF93-A99A66E97476 |
|
DOI |
https://doi.org/10.5281/zenodo.6038528 |
|
persistent identifier |
https://treatment.plazi.org/id/8B6387AE-166C-FF99-EB8D-FA886C839C6C |
|
treatment provided by |
Plazi |
|
scientific name |
Canthyporus reebae |
| status |
sp. nov. |
Canthyporus reebae View in CoL sp. nov.
Figures 1–9 View FIGURES 1 – 2 View FIGURES 3 – 6 View FIGURES 7 – 9 .
Type locality. Madagascar, former province of Fianarantsoa, south-eastern part of the Itremo mountain range, ca. 3.5 km South of Ambalamanaka , large densely vegetated pond close to N7 road, GPS coordinates S20°46' E47°10', altitude 1,700 m GoogleMaps .
Type material. Holotype (♂): " Madagascar . ex-Prov. Fianarantsoa. ca. 3.5 km S Ambalamanaka. S20°46' E47°10'. 19.V.2016. Manuel & Ramahandrison leg.", " Alt. 1,700 m. Near end of small stream, on margins of large pond with Nymphaeaceae and peat bog with Sphagnum and Drosera . Close to N7 road, south-eastern part of Itremo mountain range " [printed], “ Holotype, Canthyporus reebae sp. nov., Manuel & Ramahandrison 2017” [red, printed] ( MNHN). GoogleMaps
Paratypes (in total 14♂, 20♀) (CMM, MNHN, BMNH, NMPC): 4♂, 8♀, same data as holotype GoogleMaps . 2♂, 1♀, " Madagascar. ex-Prov. Fianarantsoa. ca. 2.5 km SSW Antanifotsy. S22°08' E46°53'. 21.V.2016. Manuel & Ramahandrison leg.", " Alt. 1,600 m. Seepage in meadow surrounded by forest. Andringitra National Park, eastern slope of massif" GoogleMaps . 1♂, 1♀, " Madagascar. ex-Prov. Fianarantsoa. ca. 2.3 km SW Antanifotsy. S22°07' E46°53'. 22.V.2016. Manuel & Ramahandrison leg.", " Alt. 1,580 m. Small ditch with clear circulating water, near trail between bridge and campground 1. Andringitra National Park, eastern slope of massif" GoogleMaps . 2♂, 3♀, " Madagascar. ex- Prov. Fianarantsoa. ca. 4.2 km SW Antanifotsy. S22°08' E46°52'. 23.V.2016. Manuel & Ramahandrison leg.", " Alt. 2,140 m. Small stream with many Cyperaceae and peaty substrate, very slowly flowing clear water, in subalpine grassland. Andringitra National Park, eastern slope of massif" GoogleMaps . 5♂, 7♀, " Madagascar. ex-Prov. Fianarantsoa. ca.
4.3 km SW Antanifotsy. S22°08' E46°53'. 23.V.2016. Manuel & Ramahandrison leg.", " Alt. 1,990 m. Grassy brooklet, very shallow, in subalpine grassland. Andringitra National Park, eastern slope of the massif". All paratypes with the respective printed red label. GoogleMaps
Description. The description refers only to the male holotype, until paragraph "aedeagus" inclusively.
Habitus ( Fig. 1 View FIGURES 1 – 2 ): Oval-oblong, with sides widely rounded, posteriorly rather strongly attenuated to narrowly rounded apex; maximum width slightly before midlength; lateral outline continuous between pronotum and elytron; dorsal surface strongly and evenly convex.
Colour ( Figs. 1 and 2 View FIGURES 1 – 2 ): Dorsal surface of head uniformly dark brown except diffusely delimited rufo-testaceous area along clypeal margin. Pronotum rufo-testaceous; disk with diffuse brown central longitudinal marking, extending from shortly behind anterior margin to posterior margin, vaguely and shortly extended transversally along posterior margin. Elytron dark brown; with rather broad diffusely delimited rufo-testaceous area along lateral margin (visible only in lateral view); within pale lateral area with narrow submarginal dark brown stripe in posterior half; near apex diffusely paler except along suture. Venter of head, pronotal and elytral epipleura, propleuron and posterior third of abdominal ventrite VI rufo-testaceous; prosternum, abdominal ventrite I and metacoxal lobes ferrugineous; rest of ventral surface black, vaguely paler along posterior margin of abdominal ventrites III–V.
Head: Anterior margin of clypeus broadly and evenly rounded, without clypeal bead or rim. Clypeal depressions broad and shallow, with small and inconspicuous punctures. Surface shining, with moderately impressed isodiametric reticulation; punctation sparse and extremely fine; without setae. Along inner eye margins with shallow longitudinal depressions and inconspicuous punctation. Antennae testaceous with narrow dark rims around distal extremity of antennomeres V–X; antennomeres V-X slightly longer than broad. Palpi entirely testaceous.
Pronotum: Rather broad; with maximum width at posterior angles. Anterior margin weakly arcuate. Posterior margin bisinuate around centre; laterally weakly curved anteriorad. Lateral margins moderately convergent and evenly arcuate from posterior to anterior angle ( Fig. 1 View FIGURES 1 – 2 ), with narrow but well-defined bead. Anterior angles acute and narrowly rounded; posterior angles rectangular and sharp. Surface shining, with weakly impressed isodiametric reticulation; extremely finely and sparsely punctate; without setae. Close to lateral margins with larger, weakly impressed punctures in shallow depression; along anterior margin with transverse row of small coarse punctures, interrupted in middle; larger but shallower punctures present along posterior margin in outer two thirds.
Elytra: Surface shining, with weakly impressed reticulation; on disk meshes slightly elongate in longitudinal direction; with extremely fine and sparse punctation; with few larger weakly impressed punctures near suture and between discal and lateral puncture rows; with very sparse short setae. Discal and lateral puncture rows discernible but rather inconspicuous. Lateral margin of elytron in lateral view almost straight behind shoulder, with narrow bead; anterior part of lateral bead visible in dorsal view. Just before apex with small, shallow depression, but without apico-lateral carina.
Ventral surface: Entirely very shining except on genae and median region of prosternum. Gula with very obsolete reticulation; genae with strongly impressed isodiametric reticulation. Antero-medial region of prosternum with surface microgranulose and with very short setae. Prosternal column (declivitous part of prosternum between procoxae) moderately ascending posteriad, with surface microgranulose, between procoxae with minute protuberance at level of anterior third of procoxae. Prosternal process weakly inclined; rather broad, oval; in proximal half laterally bordered by narrow bead; surface almost flat, near margins microgranulose, in centre smooth, impunctate; apex broadly rounded, located in impression in metaventrite. Metaventrite and metepisternum with obsolete reticulation, impunctate. Metaventrite without longitudinal lines adjacent to midline. Elytral epipleura with obsolete and irregular reticulation, impunctate; at shoulders as broad as mesotibia distally, narrowing rather abruptly between anterior third and midlength of elytra, then continuing as narrow ridge to apex. Metacoxal plates with weakly impressed longitudinally elongate reticulation, impunctate. Metacoxal lines finely impressed, subparallel (very weakly divergent anteriad) ( Fig. 2 View FIGURES 1 – 2 ); almost reaching posterior margin of metaventrite. Between metacoxal lines, surface with weakly impressed isodiametric reticulation, with few micropunctures; midline strongly impressed. Metacoxal lobes rather broad, rounded and strongly diverging laterally. Abdominal ventrites I-III fused; junction between ventrites I and II visible; junction between ventrites II and III visible except laterally and medially. Surface of ventrites with obsolete reticulation, almost impunctate; on ventrites III-VI with sparse short setae. Reticulation on ventrites I and II longitudinally elongate, on ventrites III-V transversally elongate, on ventrite VI isodiametric except along anterior margin. Ventrite VI with very shallow transverse depression.
Legs ( Fig. 2 View FIGURES 1 – 2 ): Entirely rufo-testaceous. Metafemur without apical row of short spines; without medio-posterior expansion. Protibia distally moderately broad, with anterior margin very slightly sinuate. Mesotibia triangular, distally very broad, with anterior and posterior margins straight. Pro- and mesotarsi pseudo-tetramerous. Pro- and mesotarsomeres I–III distinctly enlarged, ventrally with few large, flat and circular suction setae. Mesotarsomere V distinctly longer than protarsomere V. Pro- and mesotarsal claws small, very narrow, weakly curved; anterior and posterior claws of all tarsi equal.
Aedeagus: Median lobe in lateral view ( Fig. 3 View FIGURES 3 – 6 ) distinctly constricted above basal piece; with rather short dorsal process located in apical half. Dorsal process straight, perpendicular to main axis of median lobe. Apical part weakly curved in dorsal direction, rather evenly narrowed to apex; apex with small dorsally-directed hook. Median lobe in ventral view as in Fig. 4 View FIGURES 3 – 6 . Parameres as in Figs. 5–6 View FIGURES 3 – 6 ; apex of parameres without setae; apex of left paramere distinctly more broadly truncated than apex of right paramere.
Females: As males, except: some females with dorsal reticulation distinctly more impressed than in males; pro- and mesotarsomeres I–III narrower than in males, without suction setae. External genitalia as in Figs. 7–8 View FIGURES 7 – 9 . Spermathecal tract as in Fig. 9 View FIGURES 7 – 9 ; spermatheca long and extended, with complex loop; sclerotisation of tubuliform ductus divided by soft, almost transparent part.
Measurements: Holotype: total length (TL) = 2.2 mm, total length without head = 1.9 mm, maximum width (MW) = 1.15 mm, ratio TL/MW = 1.93. Paratypes: TL = 2.1–2.3 mm (2.2 ± 0.05), TL without head = 1.8–2.0 mm (1.9 ± 0.05), MW = 1.1–1.2 mm (1.15 ± 0.05), TL/MW = 1.85–2.0 (1.95 ± 0.05).
Variation: Paratypes vary slightly in habitus, some specimens being somewhat narrower or wider than the holotype and some being posteriorly a bit more strongly attenuated. All specimens have the pronotum much lighter than the head and elytra and with a conspicuous dark central marking, the latter of slightly variable extension and colour intensity. The extension of the pale area near elytral apex is also slightly variable. The discal puncture row on elytra is always rather inconspicuous but its punctures vary somewhat in size and depth among specimens. In all females from the type locality and in some females collected in the Andringitra massif, the dorsal reticulation is distinctly more impressed than in males; however, in some females from Andringitra it is more or less as in males.
Differential diagnosis. Within the genus Canthyporus , C. reebae sp. nov. is a member of the C. hottentottus - group. Species of this group are characterised (in males) by the median lobe with a dorsal process and the parameres without apical incision and apical setae, and (in females) by an extended spermathecal tract with a loop ( Biström & Nilsson 2006).
Canthyporus pauliani View in CoL , the only species of the genus Canthyporus View in CoL known from Madagascar prior to this study, also belongs to the C. hottentottus View in CoL -group. Detailed descriptions of this species, with illustrations of habitus and male genitalia, can be found in Guignot (1951) and in Biström & Nilsson (2006) (the median lobe in lateral view is also reproduced in Guignot 1959). In addition, the senior author (MM) has recently studied the type material of C. pauliani View in CoL (one male and one female syntypes; MNHN; specimen data in Biström & Nilsson 2006). Canthyporus reebae View in CoL sp. nov. differs from C. pauliani View in CoL in many respects:
- size larger (according to Biström & Nilsson 2006 body length of C. pauliani View in CoL 1.92–1.94 mm);
- habitus wider, with the sides more strongly rounded and the apex more narrowly rounded, with the maximum width more anterior and the dorsal surface more strongly and evenly convex ( C. pauliani with body sides subparallel and dorsum rather flat);
- colour much lighter on pronotum (in C. pauliani , pronotum entirely dark brown except rufo-testaceous lateral margins), and pale areas on elytral sides and apical region more extended;
- clypeal margin more rounded (subtruncated in middle in C. pauliani );
- antennomeres VI-XI much lighter (entirely darkened in C. pauliani );
- pronotum sides less strongly arcuate; lateral outline continuous between pronotum and elytron (distinctly discontinuous in C. pauliani );
- on head, pronotum and elytra, surface more shiny, reticulation much less impressed and punctation finer; discal puncture row on elytra much more inconspicuous;
- colour of ventral surface darker (areas that are black in C. reebae sp. nov. are ferrugineous in C. pauliani );
- ventral surface more shiny, with reticulation more weakly impressed;
- prosternal process broader;
- metacoxal lines slightly more weakly divergent;
- pro- and mesotarsi less strongly broadened;
- protarsal claws narrower and straighter (strongly curved in C. pauliani );
- different shape of the median lobe of aedeagus (see Fig. 2 View FIGURES 1 – 2 in Guignot 1951, and Figs. 131 and 132 in Biström & Nilsson 2006), particularly in lateral view.
Despite these numerous morphological differences, C. reebae sp. nov. and C. pauliani have in common a dorsal process of the median lobe which in lateral view is straight, rather thin, and perpendicular with respect to the main part of the median lobe. Within the C. hottentottus -group, according to the figures in Biström & Nilsson (2006), only three additional species share this particular configuration of the median lobe: C. alpestris Guignot, 1936 (from Kilimanjaro, Tanzania), C. petulans Guignot, 1951 (from South Africa and Lesotho), and C. loeffleri Wewalka, 1981 (from Ethiopia). According to published descriptions, C. loeffleri has many similarities with C. reebae sp. nov. (notably in size, habitus, colour, reticulation and punctation), but the two species differ at least by the colour of the pronotum (black in C. loeffleri ) and antennomeres V-XI (brownish in C. loeffleri ) and by the shape of the median lobe of aedeagus. Type material of C. alpestris (the male holotype and the single male paratype) and C. petulans Guignot, 1951 (two paratypes, one male and one female), and several non-type specimens of the latter species, have been studied by the senior author (MNHN; specimen data in Biström & Nilsson 2006). Canthyporus alpestris differs from C. reebae sp. nov. mainly by: antennomeres V-X darker and slightly more elongated; much more strongly impressed dorsal reticulation; slightly larger punctures on pronotum and elytra; more strongly impressed discal and lateral puncture rows; lateral margins of pronotum less strongly converging; ventral surface paler; and shape of the median lobe. Canthyporus petulans is similar in habitus to the species described here, but is otherwise very distinct (much larger, with central marking of pronotum more contrasted, with elytra paler, with dorsal reticulation more impressed and dorsal and ventral punctures much larger and coarser, and different shape of the median lobe).
Habitat. All specimens of the type series were found in May 2016, in two distinct areas: the south-eastern part of the Itremo mountain range (a single collecting site which is the type locality of the new species) and the eastern slope of the Andringitra massif (four collecting sites). The altitude range of the five collecting sites was 1,580 m – 2,140 m. The type locality is a large pond with dense vegetation of Nymphaeaceae and helophytes, bordered by a peat bog with Sphagnum (indicative of high acidity) and Drosera madagascariensis ( Fig. 10 View FIGURES 10 – 15 ). Here, specimens of C. reebae sp. nov. were taken at small depth, not everywhere along margins but close to a small stream that was ending into the pond (not visible on Fig. 10 View FIGURES 10 – 15 ). In Andringitra, specimens were collected below tree line at two sites: seepage in meadow surrounded by forest ( Figs. 11 & 12 View FIGURES 10 – 15 ); small, grassy slowly-flowing ditch, exposed to sun but close to forest edge ( Fig. 13 View FIGURES 10 – 15 ). The species was also found above tree line, in two very slowly-flowing streams in subalpine wet grasslands. The first one was about 50 cm deep, with peaty substratum and many Cyperaceae helophytes (locality not illustrated). In the second one, C. reebae specimens were more abundant; it was a much smaller and shallower stream with dense short grass ( Figs. 14 & 15 View FIGURES 10 – 15 ). Common features of all five collecting sites include: high altitude, open environment, clear water, very slow water circulation, substratum rich in organic matter (mineral substratum invisible), small depth, and dense marginal vegetation of Poaceae and/or Cyperaceae . Within the study area, the species was never encountered in purely stagnant water bodies or in streams and rivers with dominant mineral substrate and/or noticeably unidirectional water flow.
Note. The type locality of C. reebae sp. nov. is located about 400 m south from the type locality of Herophydrus travniceki Šťastný, 2012 .
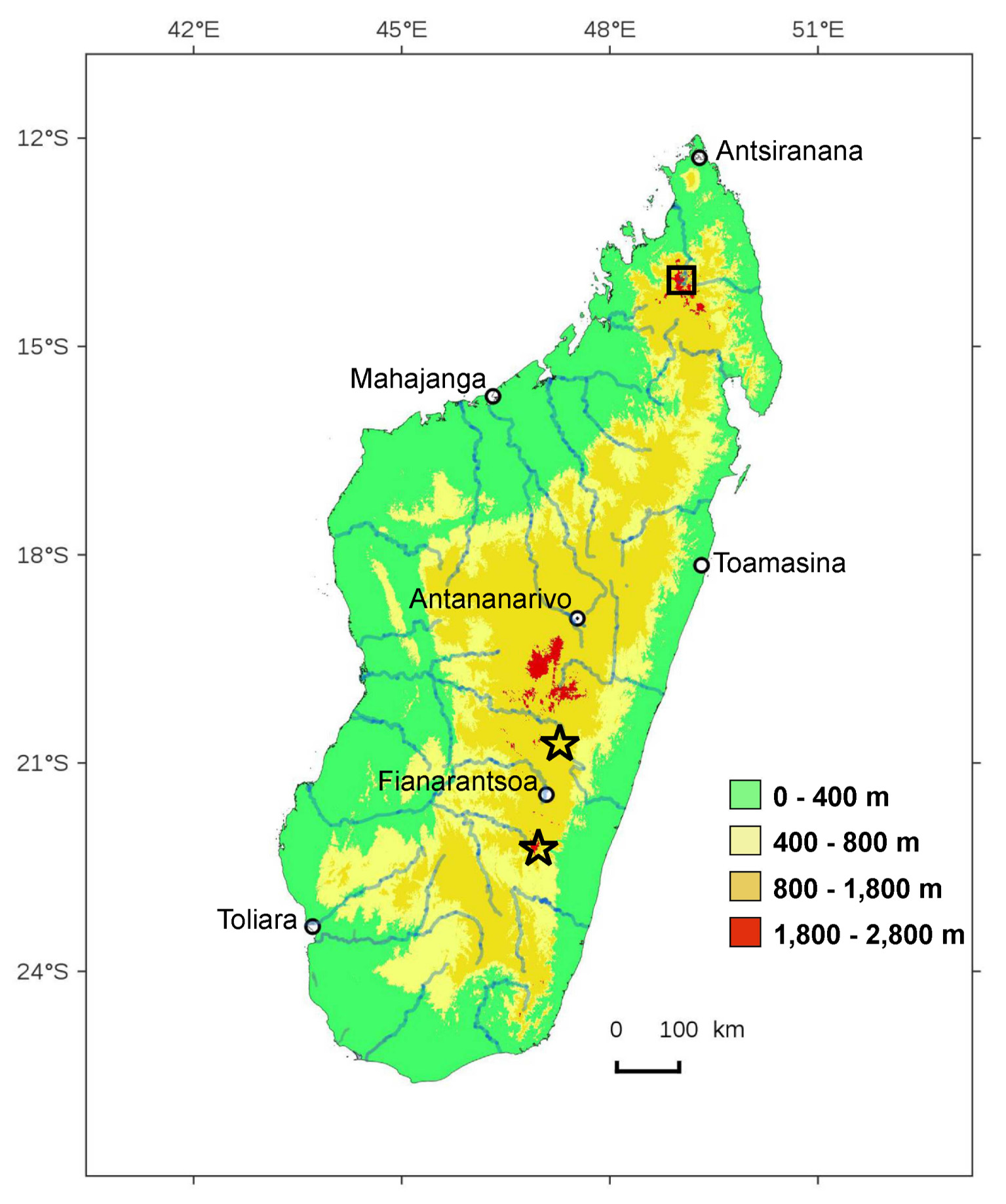
Distribution ( Fig. 16 View FIGURE 16 ). So far only known from the south-eastern part of the Itremo mountain range and from the Andringitra massif, in central eastern Madagascar, at altitudes between 1,580 and 2,140 m.
Derivatio nominis. This species is dedicated to Catherine Reeb, who organised and supervised the RicciaTeam expedition in May–June 2016, during which the specimens were collected. The species name is a noun in the genitive singular.




| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Hydroporinae |
|
Genus |