
Phrikoceros mopsus ( Marcus, 1952 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.202319 |
|
publication LSID |
lsid:zoobank.org:pub:B82E2326-417A-4CC4-8F83-E34C2C29344E |
|
DOI |
https://doi.org/10.5281/zenodo.6183437 |
|
persistent identifier |
https://treatment.plazi.org/id/8C5F87DF-8C68-FFFE-FF67-FACF8245FE24 |
|
treatment provided by |
Plazi |
|
scientific name |
Phrikoceros mopsus ( Marcus, 1952 ) |
| status |
|
Phrikoceros mopsus ( Marcus, 1952) View in CoL
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Material examined. Voucher specimen collected by M.J. Albano, 14 January 2008, Club Náutico de Mar del Plata MACN-In 38205; on mussel and ascidians, 1–2 m deep. Nine specimens, 19 February 2008, and one specimen, 9 May 2008, North Breakwater of the Mar del Plata harbour.
Morphology. Body elongate, obovate, distally rounded; approximately 45 mm long and 23 mm wide, delicate and frail in appearance. Reddish brown dorsal surface. Absence of pigment defines many whitish oval blotches regularly distributed on the dorsal surface. A dark brown mid dorsal longitudinal stripe runs along the body covering the body bulge; with a narrow delicate black rim around the body margin (figures 1A–1C). Ventral colour is paler than on the dorsal surface, with the same markings and black marginal rim (figure 1E). The marginal tentacles are dark brown, small, pointed flaps, 15 mm long, directed forwards (figures 1B). Numerous tentacular eyes present, scattered on the dorsal and ventral surface of the tentacles, especially near the fore margin. The cerebral eyes form a single elongated inverted-heart-shaped cluster located 25 mm from the fore margin, in a clear area above the cerebral ganglion region. With no frontal, marginal or ventral eyes spots.
The pharynx is 11.5 mm long and 5.5 mm in its widest part, ruffled with five short folds and laying close behind the brain (figure 1E). The mouth opens in the middle of the pharynx cavity. The main intestine opens distally in the pharynx cavity and runs dorsally and backwards along the body.
A small rounded, whitish ventral sucker locates at about mid body.
Dorsal body wall 6–7 µm high. The ciliated cellular epidermis has no rhabdites but with only granular pigmentation. Beneath the epidermis a frail layer of circular muscle fibres is present, underlain by a longitudinal muscle layer.
Ventral body wall 10–11 µm high. The ciliated cellular epidermis has no rhabdites but only some scattered granular pigmentation. Beneath the well-defined basement membrane, there is a thin layer of circular muscle fibres underlain by a layer of longitudinal muscle fibres.
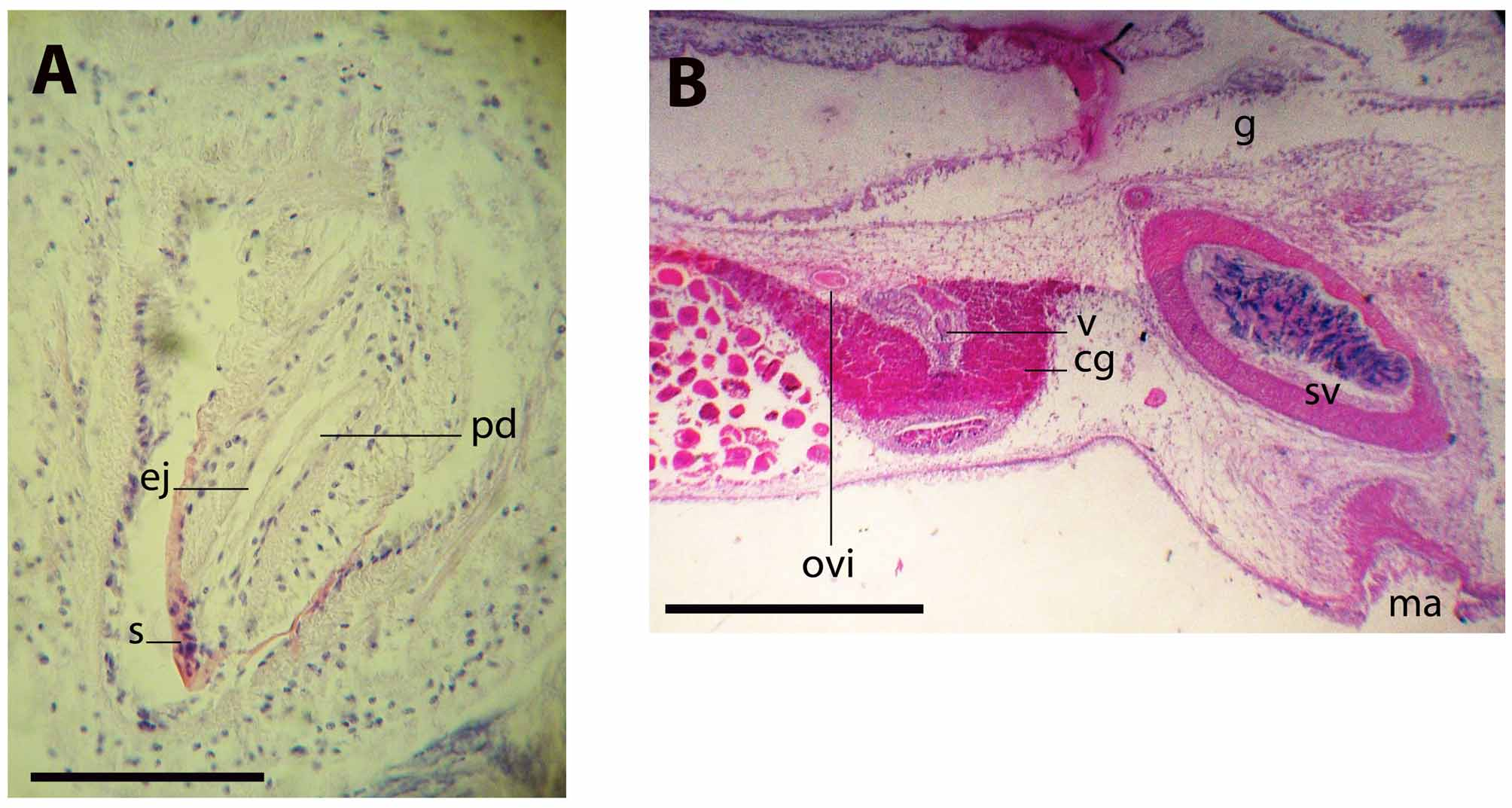
Ovaries situated dorsally, short behind the female gonopore. The uteri are well developed (figure 1D). The testes are ventral and distributed distally and to the sides of the uteri. The vasa deferentia run dorsal to the ovaries and ventral to the main intestine. They expand distally to form conspicuous spermiducal bulbs. The single copulatory system lies immediately behind the pharynx; it consists of a true seminal vesicle, free prostatic vesicle and a stylet (figure 2). Distally the spermiducal bulbs enter separately, though close to each other, the elongate seminal vesicle, which is 1360 µm long, 440 µm high, with a strong muscular wall. The rounded prostatic vesicle placed ventrally to the seminal vesicle. The prostatic vesicle is half as long the seminal vesicle; its muscular wall is 12 µm high, with a smooth glandular lining. The ejaculatory duct is underlain by a 10 µm high circular muscle layer, which follows a tortuous trajectory before entering the penis papilla and stylet (figure 3A). The prostatic duct is short, straight and joins the ejaculatory duct distally at the tip of the stylet. The stylet is conical, curved, 404 µm long; four times longer than wide and directed backwards. The penis sheath and narrow male atrium are ciliated; the male gonopore opens 19 mm behind the fore margin.
Uterine canals paired, without ramification. The oviducts enter the vagina separately from its posterior end. From this point, the vagina runs backwards and curves ventrally, leading to the cement pouch, which is a rounded expansion lined with a tall glandular epithelium (figure 3B). The vagina continues ventrally and opens to a narrow female atrium and to the exterior via the female gonopore, which opens short behind the male atrium. Aside from the cement pouch, the vagina is completely ciliated. The vagina is underlain by an apparent muscular layer of circular fibres immersed in a compact eosinophilic glandular tissue.
Despite the morphological homogeneity of the microanatomy of the examined specimens, some variation was detected. Although the colour pattern remains constant, the ground colour could vary from reddish-brown to dark brown, almost black (figure 1A, 1C). The body form varies from elongated, with rounded fore and rear margins and almost parallel lateral sides (figure 1A) to oval, with a broad fore margin, narrowing distally (figure 1C).
Some specimens were maintained in fish tanks in the laboratory where, after five days, some fertilised eggs were deposited in an 8 by 11 mm laminar sheet, adhered to the glass wall of the tank (figure 1F).
Taxonomical discussion. Marcus (1952) described Pseudoceros mopsus from the coast of Brazil. Quiroga et al. (2004) transferred the species into genus Phrikoceros Newman & Cannon, 1996 , characterized by the presence of a single male reproductive apparatus, deep marginal ruffling of the body, simple pharyngeal folds and clustered dorsal and ventral pseudotentacular eyes. Other than P. mopsus , five species of Phrikoceros were described from eastern Australia and Papua New Guinea. Although the original description of Marcus (1952) is based on a subadult specimen (not fully developed female reproductive system), the presence of marginal ruffling of the body, simple pharyngeal folds and clustered dorsal and ventral pseudotentacular eyes, as well as a single male copulatory organ with the ejaculatory and prostatic duct joining at the tip of the penis stylet characterize the species collected from Mar del Plata harbour as pertaining to Phrikoceros mopsus .
Quiroga et al. (2004) did not include any description of their examined material. We find the Mar del Plata specimens differing slightly from the original description of Marcus (1952), from the described specimen from Puerto Piramides ( Brusa et al. 2009) and from the diagnosis of the genus given by Newman and Cannon (1996) for the genus Phrikoceros .
Our specimens are large animals ranging between 43.5 and 47.5 mm long; not moderate sized as stated in the genus diagnosis given by Newman and Cannon (1996, p.1429).
Although the species belongs to the Colour Group 4 ( Newman & Cannon 1994, p. 208), this colour pattern differs slightly from the description of Marcus (1952, p. 91). Marcus’ specimens were characterized by a pattern of small sized spots covering the dorso-median region, becoming larger spots, partially fused toward the margins. The specimens from Mar del Plata had oval blotches, more or less the same size, and decreasing in number from the centre to the marginal region, without any evidence of fusion among spots. The specimen described by Brusa et al. (2009, p. 52) had a short longitudinal black line, anterior to the cerebral eye cluster that did not reached the anterior end of the body. This condition was not observed in any specimen from Mar del Plata.
Newman and Cannon (1996, p. 1429) also described the tentacles of Phrikoceros as two deep ruffles or several lateral ruffles. In our material, the morphology of the tentacles is more consistent with the morphology described by the same authors for genus Pseudobiceros Faubel, 1984 ( Newman & Cannon 1994, p. 209, figs. 3C & 3F), i.e., conspicuous ear-like pointed structures ( Marcus 1952, p. 92, fig. 173).
The cerebral eye cluster is on a clear area dorsal to the brain, but it is not small and rounded ( Newman & Cannon 1996, p. 1431, figs. 8 & 9; Marcus 1952, p. 92, fig. 173); they look as two elongated clusters fusing toward their fore part.
The pharynx of the specimens from Brazil and Mar del Plata are larger (0.25 of the body length) as compared to the description of genus Phrikoceros by Newman and Cannon (1996) (0.14–0.2 of the body length), and completely devoid of pigment, in opposition to the observations of Marcus (1952, p. 91).
Marcus (1952) described that the ejaculatory duct and the prostatic duct join each other at the base of the penis papilla. The specimens from Argentina are instead consistent with the description of genus Phrikoceros by Newman and Cannon (1996, p. 1429) in that the ejaculatory and prostatic ducts join at the tip of the penis stylet.
The sclerotised penis stylet of the specimens from Mar del Plata was neither a short, wide and straight stylet ( Newman & Cannon 1996, p. 1429), nor had an undulated margin and a terminal process ( Marcus 1952, p.92), but is elongated and curved, without any unusual formation.
The presence of spermiducal bulbs was not mentioned in Marcus’ description, maybe because his specimens were not fully developed ( Marcus 1952, p. 93).
Despite the noticed differences regarding the form of the body and the structure of the male copulatory system, the studied specimens agree with the specific features of Phrikoceros mopsus ( Marcus, 1952) and confirm its generic allocation with genus Phrikoceros .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Cotylea |
|
Family |
|
|
Genus |