
Dendropsophus juliani, Moravec, Ji Ř Í, Aparicio, James & Köhler, Jörn, 2006
|
publication ID |
https://doi.org/10.5281/zenodo.174129 |
|
DOI |
https://doi.org/10.5281/zenodo.5665978 |
|
persistent identifier |
https://treatment.plazi.org/id/8D1087CE-FF35-1A29-FEEB-B31DFB7EFD95 |
|
treatment provided by |
Plazi |
|
scientific name |
Dendropsophus juliani |
| status |
sp. nov. |
Dendropsophus juliani View in CoL View at ENA sp. nov.
Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Hyla cruzi— Köhler, 2003:381.
Dendropsophus sp.— Moravec and Aparicio, 2006:in press.
Holotype. CBF 5923, adult male, from the vicinity of the settlement of Barracón on the road from Cobija to Riberalta, 160 m a.s.l., 11°33’ S, 66°56’ W, Provincia Madre de Dios, Departamento Pando, Bolivia, collected on 26 January 2006 by Jiří Moravec.
Paratypes. CBF 5924–5927, NMP 6 V View Materials 72799/1–3, HLMDRA–3051 adult males, same locality as the holotype, collected on 26–28 January 2006 by Jiří Moravec.
Diagnosis. A small species of Dendropsophus distinguished by the following combination of characters: (1) small size, SVL 19.7–21.2 mm in males, head slightly wider than body; (2) snout short, acutely rounded in dorsal view, truncate and slightly inclined posteroventrally in lateral view; (3) canthus rostralis distinct, rounded in crosssection; loreal region slightly concave; (4) tympanum barely evident, round, about one third of eye length; tympanic annulus indistinct; supratympanic fold weakly developed; (5) vomerine odontophores prominent, separated medially, between choanae; (6) skin on dorsal surfaces smooth, with minute scattered tubercles; (7) tarsal fold and tubercles on outer edge of tarsus absent; ulnar folds and tubercles absent; (8) axillary membrane extensively developed; (9) fingers about one third webbed; toes about three fourths webbed; (10) bifid distal subarticular tubercle under fourth finger; (11) pectoral glands lacking; (12) generally, colouration of lorealtympanic region and flanks sharply contrasting with colouration of dorsum; (13) in life, dorsum pale yellowishtan to reddishtan at night and tan to reddishbrown by day, with inconspicuous dark brown markings; head greenish laterally at night and pale brown to dark brown by day; flanks pink at night and pale brown to dark brown by day; (14) in life, throat yellowishgreen to bluishgreen at night and yellowishgreen by day; belly light yellow, translucent greenish in posterior part; ventral surfaces of thighs translucent greenishyellow; (15) in life, iris periphery light silver brown with dark brown vermiculation, inner iris dark brown; bones white; (16) advertisement call composed of a series of very short notes rapidly repeated at a rate of about 10–12 notes/second, each note containing 3–6 pulses.
Comparisons. Dendropsophus juliani shares most characters with species currently placed in the D. microcephalus group (sensu Faivovich et al., 2005). Superficially, the new species is most similar to D. coffea (Köhler, Jungfer and Reichle) , D. cruzi (Pombal and Bastos) , D. meridianus (B. Lutz) and D. minusculus (Rivero) by sharing a tan dorsal colouration sharply contrasting with the flanks. However, D. juliani mainly differs from these four species by the advertisement call characteristics and by exhibiting a greenish vocal sac in life ( Murphy, 1997; Pombal and Bastos, 1998; Köhler et al., 2005). Three other small species, D. nanus (Boulenger) , D. sanborni (Schmidt) and D. walfordi (Bokermann) , differ from D. juliani by having numerous thin brown lines on a yellowish dorsum, a longer, more pointed snout and a different advertisement call ( Martins and Jim, 2003). Superficially, individuals of D. juliani might be similar to D. leali (Bokermann) in size, dorsal colouration and skin texture. However, D. leali lacks greenish ventral colours in life as well as the dark brown lateral stripe. In addition, it emits a different advertisement call ( Köhler and Lötters, 2001). Dendropsophus joannae (Köhler and Lötters) from northern Bolivia is distinguished from D. juliani mainly by having tuberculate dorsal skin, a red inner iris in life, smaller size, and a different advertisement call ( Köhler and Lötters, 2001). Dendropsophus bipunctatus (Spix) and D. studerae (CarvalhoeSilva, CarvalhoeSilva and Izeckson) from northeastern Brazil mainly differ from D. juliani by having a loreal region with numerous white blotches (CarvalhoeSilva et al., 2003).
Other small species of Dendropsophus known from the Amazonian region and either associated with the D. microcephalus group, the weakly defined D. minimus group or not associated with any particular species group (see Faivovich et al., 2005) include: D. aperomeus (Duellman) , D. haraldschultzi (Bokermann) , D. mathiassoni (Cochran and Goin) , D. microcephalus (Cope) (including the subspecific form D. m. miserus (Werner)), D. minimus (Ahl) , D. miyatai (Vigle and GoberdhanVigle) , D. rhodopeplus (Günther) , D. riveroi (Cochran and Goin) and D. tintinnabulum (Melin) . All of these species differ from D. juliani by morphological and/or bioacoustical characters: D. aperomeus exhibits a white supracloacal stripe (Duellman, 1982), lacking in D. juliani ; D. haraldschultzi has a more slender body with thin longitudinal lines on dorsum ( Rodríguez and Duellman, 1994); D. mathiassoni has a dorsum without any pattern and dorsolateral lymphatic sacs visible through the skin ( Cochran and Goin, 1970); males of D. microcephalus are larger in size and exhibit a yellow vocal sac in life ( Savage, 2002); D. minimus has a concealed tympanum ( Köhler et al., 2005); D. miyatai exhibits red markings on dorsum and a pink venter ( Rodríguez and Duellman, 1994); D. rhodopeplus has a yellow dorsum with red markings and a red lateral stripe (Duellman, 1974); D. riveroi lacks contrasting colouration and has a different advertisement call ( Köhler et al., 2005); and D. tintinnabulum has a highpitched belllike advertisement call ( Lutz, 1973).
Species in the D. rubicundulus clade of the D. microcephalus group (sensu Faivovich et al., 2005) differ from the new species by having a green dorsum in life that changes to pinkish or violet in preservative, a regular dorsal stripe pattern and longer, more pointed snouts ( Napoli and Caramaschi, 1998; 1999). From species of the D. decipiens clade (sensu Faivovich et al., 2005), D. juliani differs by colour pattern and advertisement call ( Lutz, 1973; CarvalhoeSilva et al., 2003).
Description of holotype. Body moderately robust; head wider than body, shorter than wide, widest below eyes; snout acutely rounded in dorsal view, truncate and slightly inclined posteroventrally in lateral view; distance from nostril to eye shorter than diameter of eye; canthus rostralis distinct, rounded; loreal region slightly concave; lips slightly flared; internarial area slightly depressed; nostrils barely protuberant, directed dorsolaterally; interorbital area flat, IOD 131.6% of ELW; eye large, strongly protuberant, its diameter about four times depth of lip below eye; tympanic membrane small, round, barely evident, its diameter about one third of eye length, separated from eye by slightly larger distance than its diameter; tympanic annulus indistinct; supratympanic fold evident but weakly developed, slightly obscuring upper edge of tympanum. Arm slender, not hypertrophied; axillary membrane extending to second third of upper arm; ulnar folds and tubercles absent; fingers of medium length, bearing small, round discs; relative length of fingers 1<2<4<3; diameter of disc on third finger about the size of tympanum; subarticular tubercles small, ovoid, distal one of fourth finger prominent, bifid; supernumerary tubercles barely evident; palmar tubercle small, flat, elliptical; prepollical tubercle large, flat, elliptical, lacking nuptial excrescences and glands; fingers about one third webbed; webbing basal between fingers one and two; webbing formula for fingers II 1 1/3— 3 III 2— 2+ IV. Legs moderately long, slender; heels overlapping when limbs flexed perpendicular to axis of body; tarsal fold and tarsal tubercles absent; toes moderately long, bearing round discs slightly smaller than those of fingers; relative length of toes 1<2<5<3<4; outer metatarsal tubercle small, obscure, round; inner metatarsal tubercle large, flat, elliptical; subarticular tubercles prominent, ovoid; supernumerary tubercles obscure; toes three fourths webbed; webbing formula for toes I 1 +—2 II 1 — 2 III 1— 2 IV 2— 1 V. Skin on dorsum, head, and dorsal surfaces of limbs smooth, with few scattered minute tubercles; skin on flanks smooth; skin on venter coarsely granular; skin on throat smooth; skin on lower surfaces of thighs slightly granular. Cloacal opening directed posteriorly at upper level of thighs; cloacal sheath consisting of two parts separated by a horizontal wrinkle, caudal part covering upper edge of cloacal opening; cloacal folds and tubercles absent. Tongue nearly round, slightly notched posteriorly, posterior and lateral margins not attached to floor of mouth; vomerine odontophores small, prominent, separated medially, between posterior halves of choanae, bearing three and two vomerine teeth (left/right); choanae mediumsized, oval; vocal slits long, extending from anterior third of lateral base of tongue to angle of jaws; vocal sac large, single, median, subgular.
In alcohol, dorsal surfaces of head, body, and limbs reddishtan with inconspicuous dark brown markings, consisting of a narrow interorbital streak, small obscure marking in prescapular region, inverted Vshaped mark in scapular region, inconspicuous mark in sacral region (posterior part of dorsum somewhat faded) and obscure small irregular flecks on shanks. Flanks with dark brown stripe, sharply outlined and contrasting against dorsum, fading in groin; groin whitish; supratympanic fold dark brown; upper lip whitish, with some scattered melanophores; loreal region dark brown; region around nostrils dark brown, tip of snout whitish; dorsal surfaces of hands and feet whitish with closely arranged melanophores (melanophores reduced on inner two fingers and toes); webbing, and posterior surfaces of thighs whitish without melanophores; cloacal sheath dark brown; lower lip whitish, posterior two thirds with scattered melanophores; throat yellowish; belly and ventral surfaces of limbs whitish.
In life, dorsal and lateral colouration differs only slightly from the preserved specimen in being somewhat darker coloured, and having more yellowish tint of the whitish parts. Fingers yellow dorsally, shanks with more distinct dark flecks forming three obscure transverse bars. Axillar region translucent fleshy white; groin translucent greenish; throat yellowishgreen; belly bright yellow, translucent greenish posteriorly; ventral surfaces of upper limbs translucent fleshy white, fingers greenishyellow ventrally; ventral surfaces of thighs, shanks, and fingers translucent greenishyellow; iris periphery light silver brown with dark brown vermiculation, inner iris dark brown; bones white.
Measurements of the holotype: SVL 20.7; HL 7.0; HW 7.5; IOD 2.5; EN 1.6; ED 2.5; ELW 1.9; TD 0.8; TL 10.3; FL 14.3.
Var i at io n. Variation of measurements and body proportions of the type specimens is given in Tab. 1 View TABLE 1 . A photograph of the paratype NMP 6 V View Materials 72799/1 is shown in Fig. 3 View FIGURE 3 ; lateral and dorsal aspects of the head of the Paratype NMP 6 V View Materials 72799/3 are depicted in Fig. 4 View FIGURE 4 . Some variation seems to be evident in distinctiveness of the tympanum. In CBF 2925 the tympanum is more evident and the tympanic annulus more distinct than in other type specimens. In contrast, the tympanum is difficult to detect in CBF 5927 and NMP 6 V View Materials 72799/2, possibly a result of fixation artifacts. In the holotype and NMP 6 V View Materials 72799/1 the supernumerary and carpal tubercles are less prominent than in the other specimens. The outer metatarsal tubercle is less pronounced in some individuals ( holotype, CBF 2925–5927, NMP 6 V View Materials 72799/1–2). The finger and toe webbing formulae vary as follows: II (1 1/2–1 2/3)—(2 2/3–3) III (2–2+)—(2–2+) IV and I (1–1 1/4)—(1+–2) II (1–1)—(2–2) III (1 –1+)—(2–2) IV (2–2)—(1–1) V. The axillary membrane extends along nearly whole upper arm in CBF 5926, NMP 6 V View Materials 72799/1–2.
Overall dorsal colouration in alcohol varies from yellowishtan to reddishbrown. Dorsal pattern varies mostly regarding presence, shape and distinctness of the dark interorbital streak, inverted Vshaped mark in scapular region and presence of other smaller markings (irregular dark small spots scattered on dorsum in CBF 5925, 5927, NMP 6 V View Materials 72799/2, and HLMDRA–3051). The sharply outlined lateral stripe of light grey colour is present in all paratypes except in CBF 5926 and NMP 6 V View Materials 72799/1. In the latter two specimens, lateral and dorsal colouration is not clearly defined and yellowish tan dorsal colouration is broken in smaller spots forming a triangle between snout and proximal margin of upper eyelids, two longitudinal dorsolateral bars extending from posterior margin of the upper eyelids to scapular area, and a pattern of irregularly scattered spots on the presacral and sacral region. The colour pattern of the spotted specimen (NMP 6 V View Materials 72799/1) is shown in Fig. 3 View FIGURE 3 , and that of a calling male (not collected) in Fig. 5 View FIGURE 5 .
Advertisement call. Seven calls of four individuals were analyzed. Air temperature during recording was 24°C. The recordings were obtained from a small chorus of males. The call ( Fig. 6 View FIGURE 6 ) consists of a long series of very short notes repeated in regular intervals. Notes are distinctly pulsed and lack any frequency modulation. The initial note of a call might be slightly longer than subsequent notes. Call energy is distributed within a narrow frequency band between 3300–4400 Hz. Numerical call parameters are as follows (range followed by mean ± standard deviation in parentheses): call duration, 656–1108 ms (847.5
207.2); notes/call, 7–14 (10.0 2.8); note duration, 11–26 ms (17.6 5.1); pulses/note, 3–6 (3.8 1.1); note repetition rate, 10.9–12.3 notes/sec (11.5 0.5); maximum call energy at 3427–4114 Hz (3773 199). Calls were repeated at an approximate rate of 8–12 calls per minute.
Distribution and ecology. Dendropsophus juliani is known from the surroundings of the small settlement of Barracón, Departamento Pando, Bolivia ( Fig. 7 View FIGURE 7 ). Furthermore, a population from the northern Departamento Santa Cruz is tentatively referred to D. juliani ( Fig. 7 View FIGURE 7 ; see Discussion). The type locality is situated in an alluvial plain of the small river Arroyo Jenichiquia. In general, the area consists of tall western Amazonian forest represented mostly by old secondary forest with many large emergent trees (e.g. Bertholletia excelsa , Dipteryx micrantha , Ficus schultesii , etc.), which were not selected for timber harvest 30–40 years ago. An important feature of the area is the presence of isolated enclaves of open or overgrown pampas associated with poorly drained soils, which penetrate the closed forest from the South and East ( Alverson et al., 2003). At the type locality, D. juliani was a relatively common species. Males called from graminoids emergent from small lakes or flooded depressions or from bushes over the water surface (ca. 5–50 cm above water). Nevertheless, single males were observed calling from bushes and young trees up to 250 cm above the ground and a chorus of males was heard calling from dense bushes and trees growing on the banks of Arroyo Jenichiquia. Calling activity started approximately 30 min. before sunset. Other hylid species found in sympatry with D. juliani include Dendropsophus leucophyllatus (Beireis) , D. minutus (Peters) , Hypsiboas boans (Linnaeus) , H. cinerascens (Spix) , H. lanciformis Cope , Phyllomedusa bicolor (Boddaert) , P. c a m b a De la Riva, Scinax ruber (Laurenti) and S. parkeri (Gaige) . Females and larvae of D. juliani are unknown.
Etymology. The specific name is a patronym for our colleague and friend Julián Faivovich in recognition of his important contributions to hylid frog systematics.





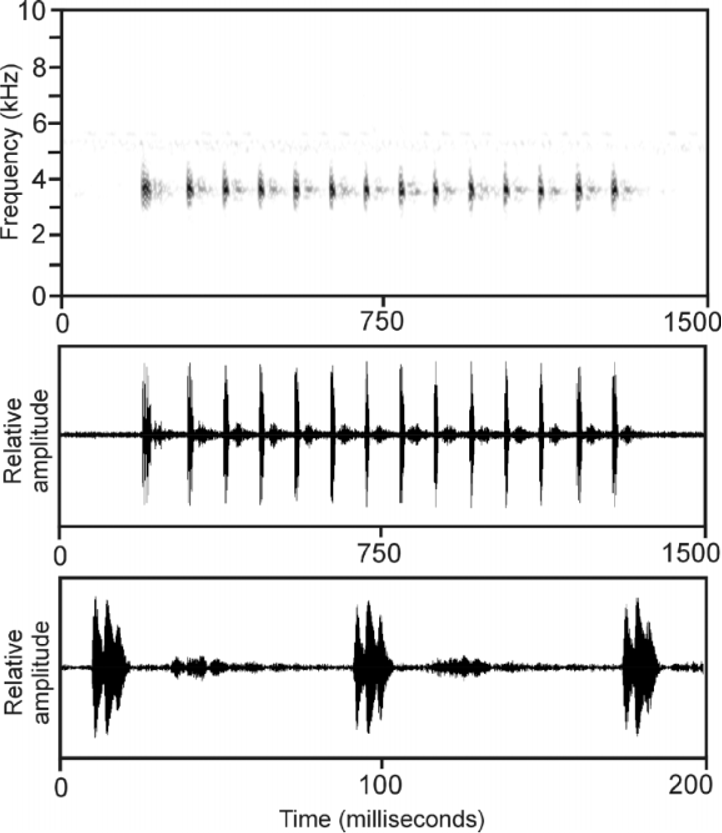
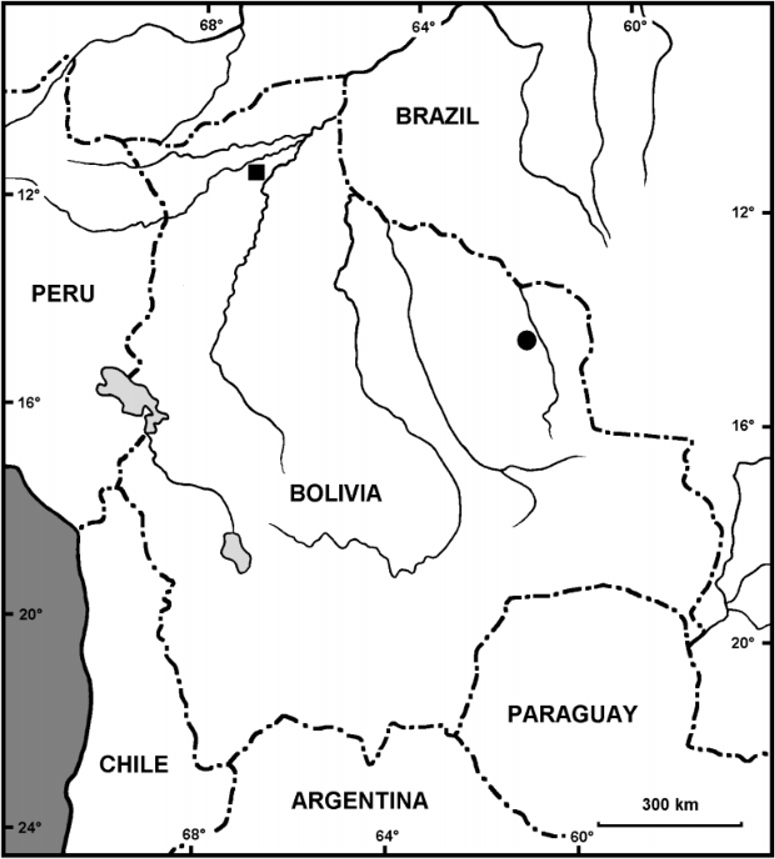
TABLE 1. Variation of measurements (in mm) and proportions of the type series of Dendropsophus juliani sp. nov. (adult males, N = 9). See text for abbreviation.
| Measurement proportions | Mean (± SD) | Range |
|---|---|---|
| SVL | 20.7 ± 0.51 | 19.7–21.2 |
| HL | 7.1 ± 0.16 | 6.9–7.4 |
| HW | 7.5±0.16 | 7.3–7.7 |
| IOD | 2.4 ± 0.08 | 2.3–2.5 |
| EN | 1.7 ± 0.05 | 1.6–1.7 |
| ED | 2.6 ± 0.09 | 2.5–2.8 |
| ELW | 1.8 ± 0.07 | 1.7–1.9 |
| TD | 0.9 ± 0.07 | 0.8–1.0 |
| TL | 10.7 ± 0.34 | 10.2–11.2 |
| FL | 14.9 ± 0.47 | 14.3–15.7 |
| HL/SVL | 0.34 | 0.33–0.35 |
| HW/SVL | 0.36 | 0.35–0.37 |
| HW/HL | 1.05 | 1.01–1.07 |
| IOD/ED | 0.93 | 0.88–1.00 |
| EN/ED | 0.63 | 0.61–0.65 |
| ED/HL | 0.37 | 0.35–0.39 |
| ED/HW | 0.35 | 0.33–0.37 |
| ELW/IOD | 0.76 | 0.72–0.79 |
| TD/ED | 0.34 | 0.31–0.38 |
| TL/SVL | 0.52 | 0.50–0.53 |
| FL/SVL | 0.72 | 0.69–0.74 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.