
Parotoplana fredigatidani, Delogu & Casu & Curini-Galletti, 2008
|
publication ID |
https://doi.org/10.1080/00222930701840696 |
|
persistent identifier |
https://treatment.plazi.org/id/8D5587EB-FFF1-AE42-19C5-FB589C304F7D |
|
treatment provided by |
Felipe |
|
scientific name |
Parotoplana fredigatidani |
| status |
|
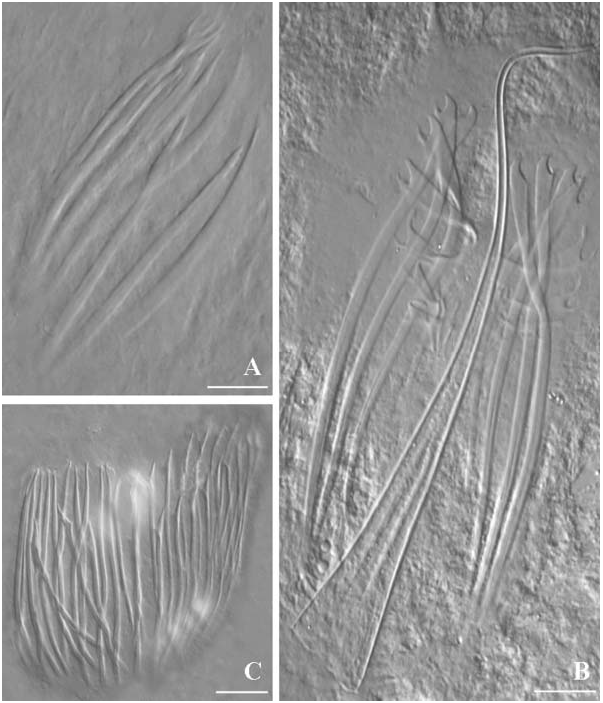
Parotoplana primitiva Ax, 1956 View in CoL ( Figures 4C View Figure 4 , 5 View Figure 5 )
Material examined
Playa el Palo, Malaga (lat. 36 ° 4393.620N, long. 4 ° 21929.060W); about 2 m deep, fine sand. Five specimens sagittally sectioned (CZM 90–94); one karyological slide, made permanent (CZM 95); two whole mounts (CZM 96–97). October 2005.
Playa les Marines, Bolaga (lat. 37 ° 11917.030N, long. 1 ° 48955.680W); about 1 m deep, medium sand. Whole mount (CZM 98). October 2005.
Tarifa, Atlantic side (lat. 36 ° 0931.060N, long. 5 ° 36933.060) about 50 cm deep, medium sand. Whole mount (CZM 99). October 2005.
Description
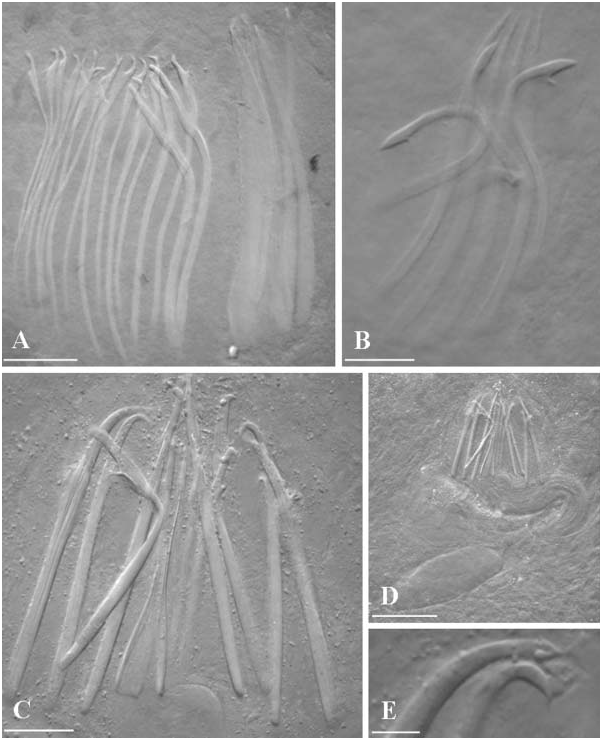
For general shape and organisation, see the original description ( Ax 1956). Male genital organs. The Spanish specimens presented two lateral rows of testes (about 11 in each row) anterior to the ovaries. Copulatory organ consisting of a sacciform seminal vesicle, distally connected to a prostatic vesicle, about 60 Mm long. Prostatic vesicle lined by a glandular, non-ciliated epithelium. It is distally connected to the sclerotized apparatus, consisting of two concentric girdles of spines. The outer girdle consists of 22–23 spines with sickle-shaped apices; dorsal spines are broader and shorter (about 40–43 Mm), spines become progressively longer and slender (about 50–52 Mm long) ventrally. The inner girdle consists of 10–13 needle-shaped spines, 48–53 Mm long.
Karyotype. With n55, and FN510. The haploid set consists of three comparatively large chromosomes, one chromosome intermediate in size, and one very small chromosome. Chromosomes are meta-submetacentric; the smallest is at the lower limit of the submetacentric class. The measurement of four spermatogonial plates yielded the following karyometrical data: Chrom. I: r.l.527.27¡0.44; c.i.545.95¡1.76 (m); Chrom. II: r.l.525.03¡1.19; c.i.541.38¡1.88 (m); Chrom. III: r.l.523.16¡0.3; c.i.535.6¡0.92 (sm); Chrom. IV: r.l.516.94¡1.09; c.i.537.19¡1.27(sm); Chrom. V: r.l.57.58¡0.03; c.i.528.97¡0.75 (sm).
Remarks
The species was described from specimens from the Atlantic coast of France (Bay of Arcachon) ( Ax 1956), the only locality where the species had been found so far. Anatomical details, as appreciable in sectioned specimens, and size and morphology of the sclerotized structures, appear practically identical in French and Spanish specimens, allowing the assumption that only a single species is concerned, whose range thus encompasses the western Mediterranean. Parotoplana primitiva widespread on the southern coast of Spain, from Bologa to Tarifa.
The karyotype of the species is quite distinctive. The chromosome number n55 is in fact the lowest known so far for the family Otoplanidae (cf. Delogu and Curini- Galletti 2007). The absence of a pair of chromosomes with respect to the basic set for the Otoplanidae (cf. Martens et al. 1989a) and the presence of a third pair of large metacentric chromosomes suggest a Robertsonian mechanism of chromosome fusion, involving two of the small chromosome pairs, that may be at the basis of the karyotype evolution of P. primitiva . Similar processes have already been extensively documented for Proseriata ( Curini-Galletti et al. 1989) .
Genus Parotoplanella Ax, 1956 View in CoL
General morphology similar to the genus Parotoplana Ax, 1956 View in CoL . Characterized by the anterior position of the ovaries, which are intercalated among testes, and by the vitellaria which, anteriorly, lie laterally to testes. Epithelium of the ciliated creeping sole with depressed nuclei. Outer epithelium of the pharynx non ciliated.
Parotoplanella heterorhabditica Lanfranchi, 1969 View in CoL
( Figure 3B View Figure 3 )
Locality. Spain, St. Pola (lat. 38 ° 11910.670N, long 0 ° 35953.260W); lower intertidal in the Otoplana -zone, medium sand.
Material examined. One whole mount (SMNH 6784).
Description
For general shape and organisation, see the original description ( Lanfranchi 1969). The specimen found had a male copulatory organ consisting of a small, globular seminal vesicle connected to a short, ovoid prostate vesicle, and a sclerotized apparatus consisting of seven spines, divided into two groups:
1) four straight, needle-shaped spines, with pointed apices, ranging from 49– 51 Mm in length;
2) three shorter spines, 38–40 Mm long, with swollen, pointed apices, and provided with well-developed, elongate subterminal teeth.
Remarks
The species was described from specimens from the Ligurian Sea (northern Tuscany, Italy) ( Lanfranchi 1969). The specimen found corresponds closely, for general arrangement of organs, and size and morphology of the sclerotized structures, to the original description.
General remarks
Previous results of the BIOIMPA campaigns concerned the composition of the genus Parotoplana in a marine protected area of the central Mediterranean (southern Apulia, Italy), where four new species were found ( Delogu and Curini-Galletti 2007). The new records bring the number of Mediterranean species to 14, of which only one ( P. primitiva ) is known to occur outside the basin (present note). Our knowledge of the composition of the genus in the Mediterranean is however far from being exhaustive, as BIOIMPA campaigns, in other sectors of the Italian coasts, have revealed a considerable number of new species, which are still in the course of being described.
At the present stage of knowledge, most species of Parotoplana appear exclusive to their type locality and surroundings, and whenever different geographic sectors are sampled, a new suite of species is found. This obviously has to do with the geographic scale of the sampling design of the BIOIMPA project, which was indeed planned with the aim to maximize the discovery of new species, with the selection of stations in different sectors of the Mediterranean. However, the congruence of the pattern found so far (new species in each area, with few or no overlap in species composition among stations) points indeed to the existence of limited distribution ranges. Ecological differences among sectors of the Mediterranean (cf. Pérès and Picard 1964), coupled with the short life cycles and lack of dispersal phases of interstitial Platyhelminthes may be at the basis of present species diversity. It is indeed reasonable to assume that interstitial organisms in general may have scales for allopatric speciation different from those of macrofaunal organisms (cf. Casu and Curini-Galletti 2006) raising questions about the global contribution to marine biodiversity of meiofaunal taxa, which is probably severely underestimated at present.
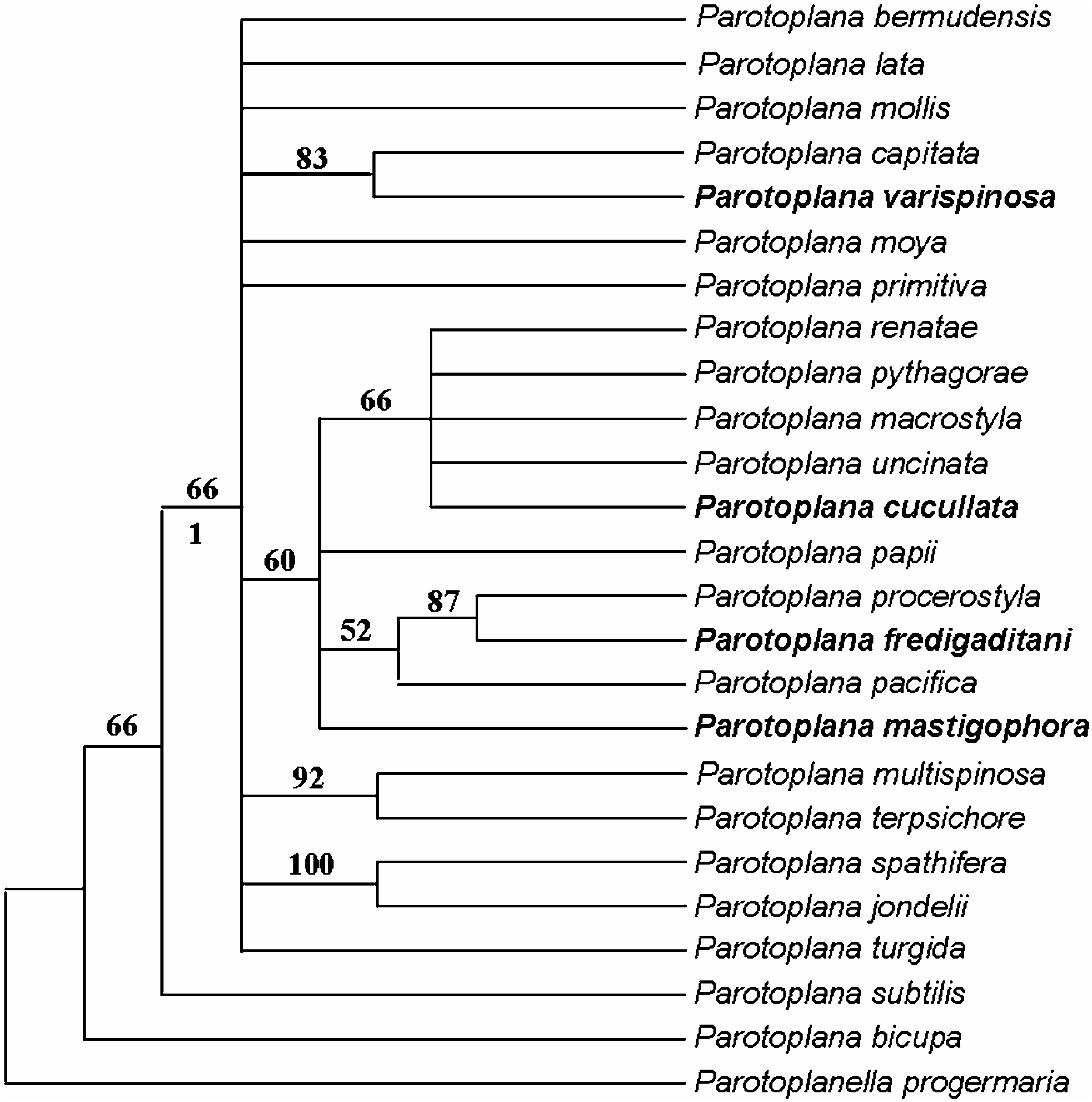
In this general context, the fauna of southern Spain is of particular interest for its geographic position. Among the species whose phylogenetic relationships could be traced, two ( P. fredigatidani sp. n. and P. cucullata sp. n.) are sister species of Mediterranean taxa, while P. varispinosa sp. n. is the sister species of an Atlantic taxon. The two species whose ranges extend beyond the coasts of southern Spain show a similar pattern, with one species known from western France ( P. primitiva ) and one from the northern Mediterranean ( Parotoplanella heterorhabditica ). Therefore, the presence, in southern Spain, of an overlap zone between Atlantic and Mediterranean faunas, well documented in macrofaunal groups, with an Atlantic component often conspicuous at the Gibraltar sill, which progressively disappears towards the eastern borders of the Alboran Sea (see, amongst others Harmelin and D’hondt 1993; Maldonado and Uriz 1995; Abello et al. 2002; Arroyo et al. 2006 and references therein), appears to apply to meiobenthic flatworms as well. Only rarely have Alboran endemics been found in macrofauna, and, when present, interpreted as Tethyan relicts of Premessinian biota ( Boury-Esnault et al. 1992). Although biased by the poor overall knowledge of south-western Europe and west African meiofauna, present data suggest that, for interstitial Platyhelminthes, the presence of species endemic to the Alboran sea may on the contrary be relevant.
Finally, there are suggestions that the same ecological constraints which apply to the Atlantic component of the macrofauna, whose eastward penetration appears in many instances limited by Palos Cape, off south-east Spain ( Abello et al. 2002), may affect meiofauna distributions as well. Species of Proseriata found in the only station sampled north of Palos Cape (St. Pola) were in most instances different from the suite of species found in the stations located in the Alboran Sea (unpl. data). In the case of the genera considered here, one species only was found at St. Pola, namely Parotoplanella heterorhabditica , a northern Mediterranean species absent from the Alboran Sea. However, hints about biogeography and patterns of biodiversity raised by the present study inevitably call for renewed and extensive work of taxonomic research.
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |
Parotoplana fredigatidani
| Delogu, Valentina, Casu, Marco & Curini-Galletti, Marco 2008 |
Parotoplanella heterorhabditica
| Lanfranchi 1969 |
Parotoplana primitiva
| Ax 1956 |
Parotoplanella
| Ax 1956 |
Parotoplana
| Ax 1956 |