
Keratoisididae, Gray, 1870
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5093.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:58B73298-6A33-4040-B898-ED3D8A69DA81 |
|
DOI |
https://doi.org/10.5281/zenodo.5910882 |
|
persistent identifier |
https://treatment.plazi.org/id/8E2A8791-8464-FFE3-5CE0-F9D5FD6EF8D9 |
|
treatment provided by |
Plazi |
|
scientific name |
Keratoisididae |
| status |
|
Details regarding Keratoisididae View in CoL clades
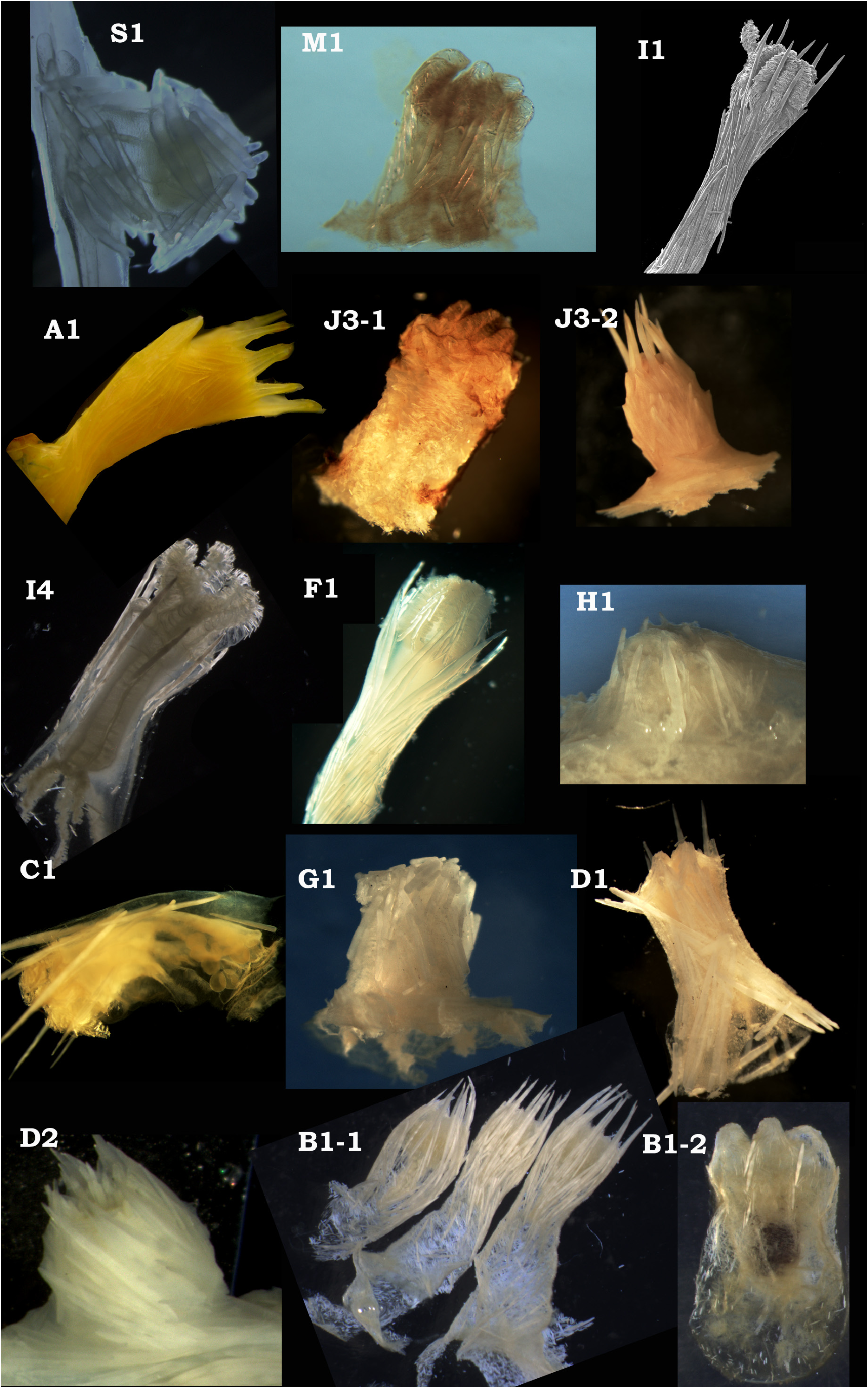
Clade 3 (S1) encompasses sequences derived from the holotype specimen of Cladarisis nouvianae Watling, 2015 , two undescribed taxa, and a specimen (USNM 75083) identified as Isidella elongata (Esper, 1788) by F.M. Bayer and collected from near the type locality in the Mediterranean Sea. Our sequence for that specimen is restricted to the mtMutS-5’ and a short 18S region. We were able to obtain a partial sequence (654bp for mtMutS- 5’: matching positions 74–727 in the GenBank sequence KC984577 View Materials ) from the holotype specimen of I. trichotoma (USNM 56715) and found it to uniquely match a portion of the haplotype of USNM 75083; however, we have been unable to replicate these results as subsequent PCR amplification attempts of the holotype specimen failed to produce an amplicon. Divergence among the 4 haplotypes of clade 3 is greater than within clade 2, with intra-clade p = 0.0005 –0.0033 for the mtMutS- 5’ region. Morphological characters of this clade include ( Table 4 View TABLE 4 ; Fig. 2 View FIGURE 2 , S 1 View FIGURE 1 ; Fig. 3 View FIGURE 3 , S 1 View FIGURE 1 ): a solid, relatively narrow-diameter axis subdivided by organic nodes; branches are sparse and originate at nodes that are moderately flexible; polyp sclerites are all rods, often thick and curved, and arranged in an irregular fashion over the surface of the polyp; on contraction, the polyp shortens and the tentacles fold over or into the apical part of the polyp.
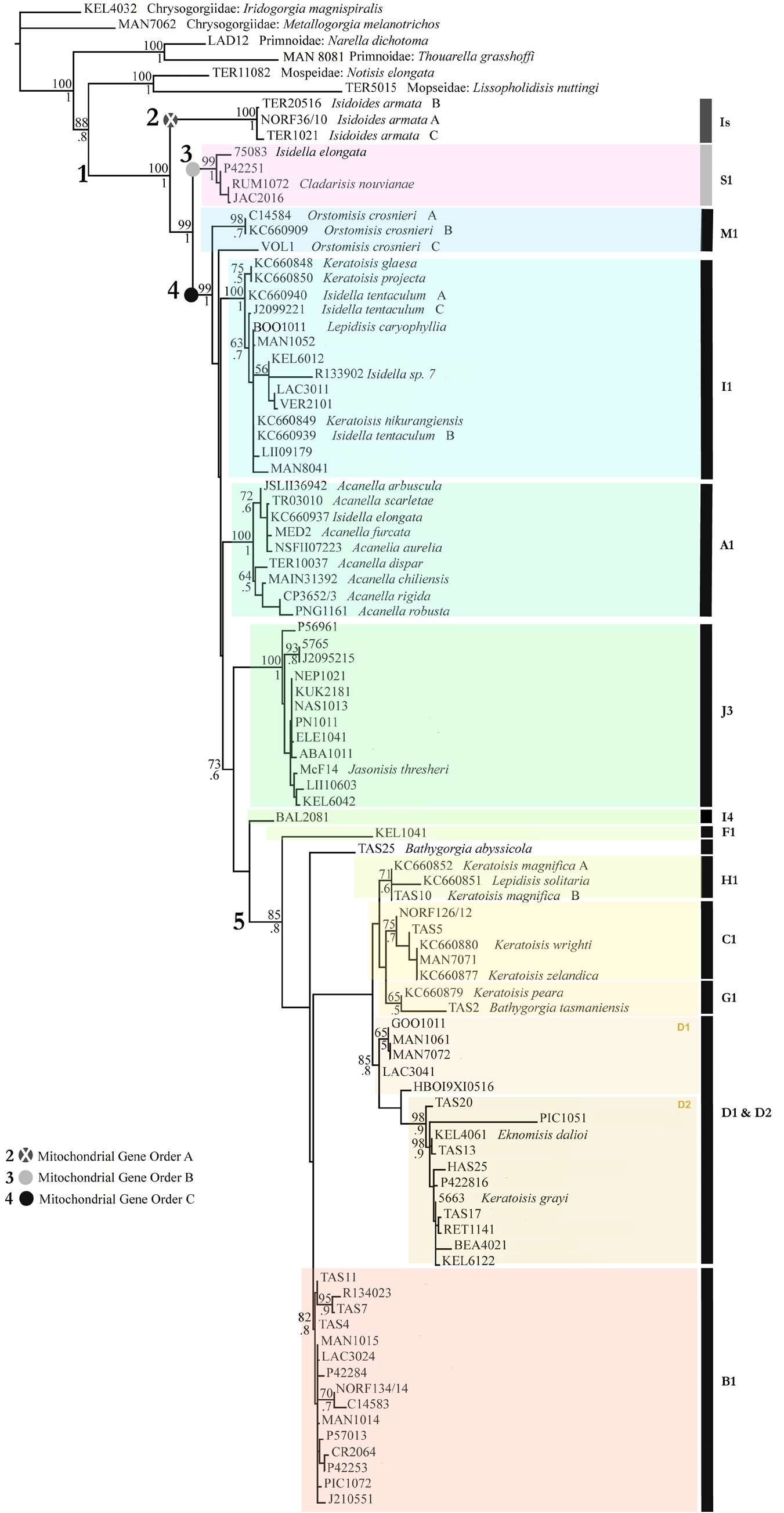
Clade 4 contains most of the diversity in the tree and most closely matches the former Isididae subfamily Keratoisidinae , which has now been elevated to the rank of family ( Heestand Saucier et al. 2021). Clade 4 is further characterized by several well-supported subclades (M1, I1, A1, J3) and a large group (5) containing subclades with varying levels of support (I4, F1, H1, C1, G1, D1 & D2, and B1) (see Fig. 1 View FIGURE 1 ); as noted earlier these subclade names follow a convention established by France (2007) based on pattern of unique indels and have been used by other investigators, so we retain them here. More genetic data will be needed to boost support for internal nodes and resolve the sister-taxon relationships among these subclades. Morphological features of these subclades are summarized in Table 4 View TABLE 4 , and selected examples of colony and polyp form are illustrated in Figs. 2 View FIGURE 2 and 3 View FIGURE 3 .
Subclade M1 comprises 2 haplotypes from two specimens both identified as Orstomisis crosnieri Bayer, 1990 and collected from near the type locality ( New Caledonia) for that species. The 18S sequence for O. crosnieri A does not differ from the 18S sequences of the syntype ( AF052916 View Materials ; Berntson et al. 2001). Both specimens were collected on the same cruise but from different stations and the sequences were derived in separate labs (A by the current authors and B from Dueñas et al. 2014); the mtMutS- 5’ haplotypes are identical.A third haplotype associated with a specimen (VOL1) identified as O. crosnieri and also collected from near the type locality is on an unsupported branch, which, if the unsupported nodes were collapsed, would result in a trichotomy with the M1 and I1 clades. Orstomisis crosnieri C differs from O. crosnieri A and B by [only] p = 0.0074 at mtMutS- 5’, and might be an undescribed Orstomisis species. Taxa in subclade M1, typified by O. crosnieri , have a solid axis; dichotomous or pseudodichotomous branching that originates at the organic nodes; polyp sclerites that are all rod-shaped; and a polyp that does not shorten on contraction but has tentacles that simply fold over the mouth area.
Subclade I1 is one of the more genetically divergent of the clade 4 subclades (along with the group at clade 5 from F1 to B1), in terms of genetic distance between haplotypes, with an intra-subclade p = 0.0005 –0.013 at mt- MutS- 5’. It includes specimens initially identified in 3 different genera ( Isidella , Keratoisis , Lepidisis ) and three different haplotypes from specimens identified as Isidella tentaculum Etnoyer, 2008 (one of which is the holotype). Sequences from three of the Keratoisis species are derived from holotype specimens, but, as discussed below, they almost certainly should not be in that genus. Members of this subclade possess a hollow, thin axis, at least in the distal parts of the colony; when present, branches originate at the nodes; polyp sclerites, when present, are rods, often in the form of needles, with long, sometimes thick needles positioned between the bases of the tentacles (referred to as intertentacular needles due to their mesenterial, or aligned with the septa, arrangement); and the polyp on contraction does not shorten but the tentacles fold over the mouth. This subclade contains Lepidisis caryophyllia Verrill, 1883 , one of the oldest known keratoisids and the type species in one of the most commonly mis-identified genera ( Watling & France 2021).
Subclade A1 contains all taxa identified as Acanella , along with a single haplotype downloaded from GenBank and identified there as Isidella elongata . Heestand Saucier et al. (2017) noted that at least some colonies identified as I. elongata could be considered to be conspecific with Acanella arbuscula . Divergence among the 8 haplotypes of subclade A1 is on par with that among the 4 haplotypes of subclade S1, with intra-subclade p = 0.0005 -0.0083 at mtMutS- 5’. Morphologically, members of this subclade are characterized by a solid axis, with branches originating at the organic nodes, usually in whorls, often of up to 5 branches; sclerites of the polyps are needles and other rods, occasionally intertentacular in position; and a polyp that does not shorten on contraction but has tentacles that simply fold over the mouth area.
Subclade J3 includes specimens identified to 5 different genera ( Isidella , Jasonisis , Keratoisis , Lepidisis , Orstomisis ) as well as unidentified taxa that are putatively new species. Specimens assigned to the genera Isidella , Keratoisis , Lepidisis and Orstomisis were provisionally identified before the genus Jasonisis was described (in 2012) and are almost certainly incorrectly identified. One of the included haplotypes in Subclade J3 comes from the type specimen for the genus Jasonisis ( J. thresheri Alderslade & McFadden, 2012 ). The Jasonisis type specimen and tentatively identified Jasonisis specimens do not share the same indel structure ( J. thresheri has 18 base pairs (6 amino acids) fewer than the other tentative Jasonisis specimens). Members of this clade show a wide diversity of colony and polyp form. Three colony forms and two polyp types are illustrated in Figs. 2 View FIGURE 2 and 3 View FIGURE 3 . Colony form ranges from simple fans with sparse branching to complex fans with multiple branches, but in the NW Atlantic at least, colonies can also take on the form perhaps best described as brambles, forming low-growing tangled arrays of mostly thin branches spreading over wide areas of the seafloor. Subclade J3 colonies may have the following characters: axis hollow, sometimes thick, solid near the colony base; branches almost always present, may originate at the nodes, immediately distal to the nodes, or, in the bramble forms, at the internodes; polyp sclerites are spindles, flat rods and scales (occasionally with a fluted margin), needles and long rods; on contraction the polyp shortens slightly, and the tentacles fold over the mouth; polyps are often covered with a layer of tissue containing many cnidae.
Subclade I4 is perhaps better referred to as “lineage I4 ”, being comprised of only a single haplotype, which we detected in 10 specimens (Supplemental Table S1 View TABLE 1 ). It was assigned a subclade name (first published in van der Ham et al. 2009) because the haplotype has a unique pattern of indels in mtMutS and igr4 relative to other keratoisidids. In our analysis, lineage I4 arises from a polytomy with subclade J3 and clade 5. I4 colony form is in the shape of a candelabra arising from a moderately elongate stalk, e.g. no branching in the lower part of the colony; the axis is solid in the lower part of the colony but hollow in the upper reaches of the branches; branching above the stalk begins as a trident, additional branches being added laterally in pairs in one plane; polyps are tall, sclerites are all rods, mostly needle-like, and sclerites are generally absent from the basal part; due to lack of proximal sclerites polyps can shorten on contraction with tentacles folding over the mouth.
Clade 5 (containing subclades F1 to B1) encompasses the greatest number of haplotypes and includes specimens identified to the genera Bathygorgia , Eknomisis , Isidella , Keratoisis , and Lepidisis . Specimens assigned to the genera Isidella and Lepidisis are almost certainly incorrectly identified and are likely undescribed species of Keratoisis , Eknomisis , or Bathygorgia , or an undescribed genus. Clade 5 includes 7 smaller clades, although some are only weakly supported, each with mtMutS -5’ haplotypes of a unique indel structure, e.g. there are 7 different patterns of indel structure within clade 5 that are not shared with other named clades of keratoisidids (clades 3 & 4). Genetic distances among mtMutS -5’ sequences within each of the 7 subclades range from p = 0.0120 – 0.0948. Included in clade 5 are the two species of the newly resurrected genus Bathygorgia , although these did not cluster together and showed a high level of divergence from each other (p = 0.10189 over the first half of mtMutS -5’, the only region we were able to sequence for B. abyssicola ); each species of Bathygorgia also has a unique mtMutS - 5’ haplotype with an indel structure not seen in any of the other specimens (n=398) analyzed for this study. The other nominal genera identified within clade 5 for the most part do not form monophyletic groups. The holotype of the monotypic genus Eknomisis is found nested within a strongly supported subclade (D2) dominated by species tentatively assigned to Keratoisis . This subclade also contains a haplotype from a specimen identified as K. grayi (an identification influenced by the collection location), which is the type species of Keratoisis and for the family Keratoisididae . Subclade G1 contains only two haplotypes, representing the holotype specimens of Bathygorgia tasmaniensis and Keratoisis peara ( Dueñas et al. 2014) . Subclade G1 is only weakly supported and the two haplotypes are divergent (as shown by the relatively long branch to B. tasmaniensis ) and also differ by a 9bp (3 amino acid) insertion in K. peara . Subclade B1 encompasses 15 different haplotypes that share the same mtMutS - 5’ indel structure and are only weakly divergent from each other. Within Clade 5 there is a wide diversity of colony types, from unbranched whips, to sparsely branched colonies, to large bushes with multiple branches. Branches (when present) are consistently internodal. Most of the haplotypes in subclade B1 come from colonies that are generally whip-like (unbranched), although one haplotype from the eastern North Pacific is associated with bushes or planar fans and two others from the central and south Pacific are branched. Conversely, most of the haplotype diversity within subclade D1&D2, is associated with colonies that are branching bushes or fans, with only a couple of unbranched forms in subclade D1. Subclade C1 is the only clade with multiple haplotypes to comprise exclusively unbranched colonies, Keratoisis wrighti (Nutting, 1910) and K. zelandica Grant, 1976 having both been described as unbranched. Clade 5 polyp sclerites range from rods and needles to flat rods and scales, usually with intertentacular (mesenterial) rods or needles. The presence of scales, or the absence of sclerites, in the proximal part of the polyp distinguishes most members of subclade B1 from the other subclades. Polyps are generally tall and thin with little to no contraction, the tentacles folding over the mouth, but in D1&D2 the genus Eknomisis is reminiscent of subclade S1, with the tentacles folding deep into the oral area, and intertentacular needles are absent, a form that is also seen in some specimens of subclades H1 and B1 ( Fig. 3 View FIGURE 3 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.