
Gerrhopilus depressiceps, Sternfeld, 1913
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4299.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:38508C24-1594-43DF-84FA-EB7DB779C371 |
|
DOI |
https://doi.org/10.5281/zenodo.6023896 |
|
persistent identifier |
https://treatment.plazi.org/id/9009D40C-1278-FFF4-FF02-F8988E7CFB8F |
|
treatment provided by |
Plazi |
|
scientific name |
Gerrhopilus depressiceps |
| status |
|
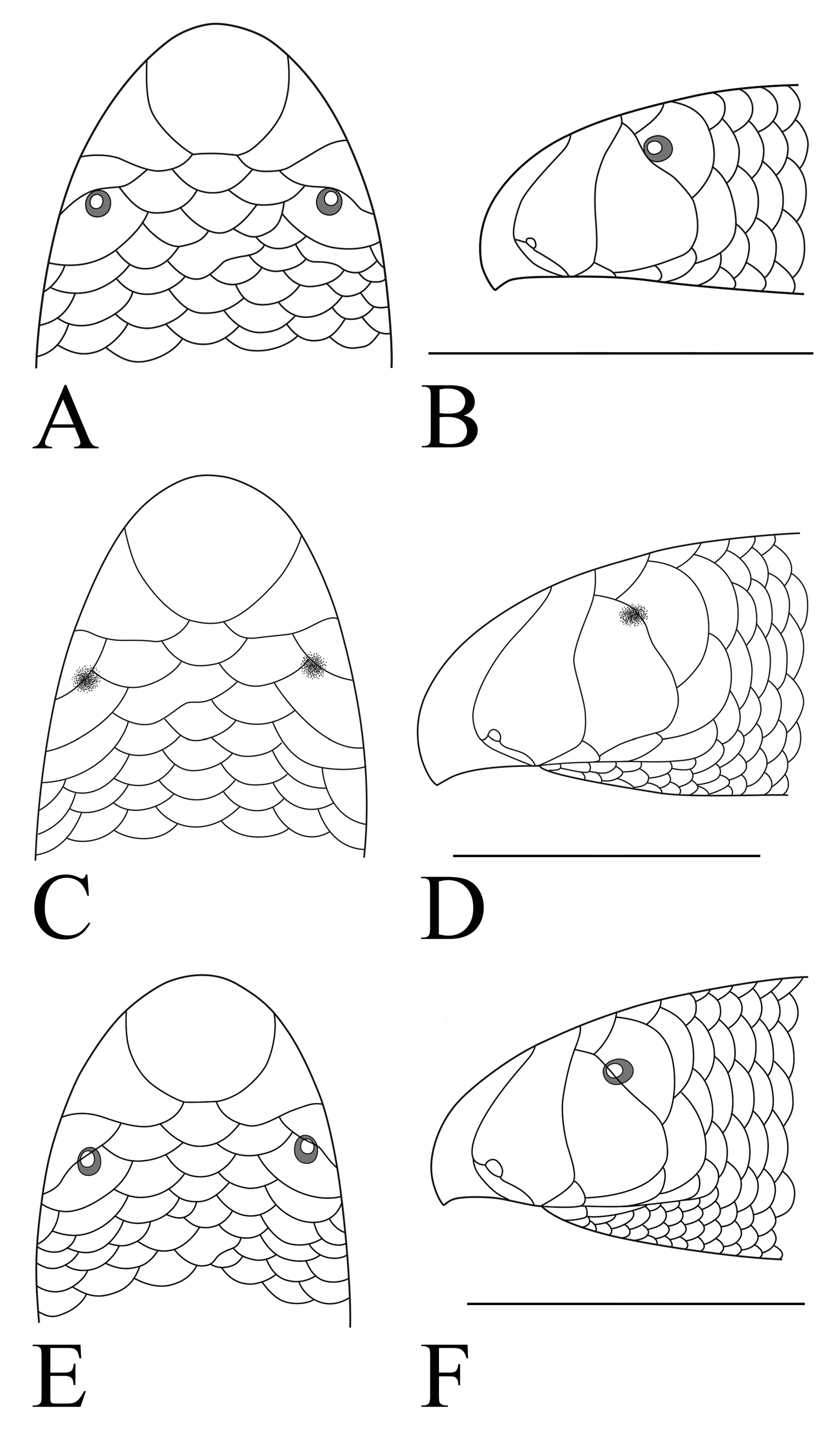
Figs. 1 View FIGURE 1 A, B
Typhlops depressiceps Sternfeld, 1913: 384 . Typhlops monochrus Vogt, 1932: 293 .
Typhlops monochrous Loveridge, 1948: 320 .
Holotype. Sex unknown, ZMB 23986, “ Neuguinea ” (= northern Papua New Guinea).
Diagnosis. This species belongs to Gerrhopilus based on the presence of head glands in the centers of the anterior head shields in addition to their anterior margins ( McDowell 1974; Wallach 1996b). A relatively robust (adult L/M = 70) species of Gerrhopilus having the unique combination of a rostrate snout with a transverse keel on the ventral margin of the rostral that extends ventral to the rictus, angle of pre-oral snout in lateral aspect horizontal, distinct pupil in the eye, preocular barely covers the anterior margin of the eye in lateral view, longitudinal scale rows 24/22/20, transverse scale rows posterior to the rostral 649–664, supralabial imbrication pattern T-V, subocular scale single, presubocular scale absent, posterior margin of rostral shallowly concave, prefrontals and supraoculars subequal in size to frontal and parietals and interparietal, subcaudal scales 27–30, L/W ratio 57–67, and tail spine oriented ventrally at an angle of 70˚ to axis of anteroventral surface of that terminal scale (and, hence, to body axis). Refer to Table 1 for additional diagnostic qualitative and quantitative features.
Comparisons with other species. Gerrhopilus depressiceps may be distinguished from all other members of this genus except G. mcdowelli in having a transverse keel on the ventral margin of the rostral, which gives the snout a beaked appearance in lateral aspect, and in having a posterior reduction of four longitudinal scale rows from head to vent. From that species G. depressiceps is distinct in its greater number of mid-dorsal scale rows (649–664 vs. 431–464 in G. mcdowelli ); greater length (188–323 mm vs. 94–199 in G. mcdowelli ); narrower body (L/W = 57–67 in G. depressiceps vs. 44–53 in G. mcdowelli ); large, transversely expanded apical spine (vs. spine small and needle-like in G. mcdowelli ); pre-oral snout oriented horizontally (vs. inclined at 30° from horizontal in G. mcdowelli ); and rostral keel pointing directly downward (vs. anteroventrally in G. mcdowelli ).
Redescription of the holotype. L = 323 mm, SVL = 313 mm, TL = 10 mm, HW = 4.5 mm, NW = 3.9 mm, SN = 2.7 mm, SW = 3.8 mm, PSN = 1.4 mm, RW = 1.95 mm, EW = 0.35 mm, W = 4.8 mm, VW = 4.5 mm, TW = 4.2 mm, L/W 67, M = 4.6 g (in preservative). Snout acutely rounded in dorsal view, rounded but with ventrally directed transverse keel in lateral view, giving rostral a beaked appearance; keel deepest medially, keratinized. Preoral snout horizontal, parallel to body axis, with rostral keel extending ventral to rictus; gular region hidden from lateral view. Rostral large (0.43 head width), oval in shape, with parallel sides dorsally, posterior border extending halfway between eye and naris, posterior margin shallowly concave; ventral surface papillose, with sides slightly diverging anteriorly, slightly concave behind keel. Nasals separated dorsally by prefrontal ( Fig. 1 View FIGURE 1 A); superior nasal large, with slightly sinuous posterior margin, crescentic dorsally, acutely rounded ventrally ( Fig. 1 View FIGURE 1 B). External naris compressed, oval shaped, oriented obliquely, close to rostral; superior nasal suture complete, extending horizontally from naris to contact rostral; inferior nasal suture complete, contacting second supralabial just posterior to latter’s contact with first supralabial. Prefrontal, frontal, supraoculars, parietals, and interparietal all subequal in size. Preocular large, triangular; larger than ocular but subequal in size to superior nasal. Ocular large, smaller than preocular, extending dorsally slightly medial to preocular, extending ventrally to ~2/3 depth of preocular, bordered posteroventrally by subocular of less than half its size. Eye with distinct round pupil and iris, situated at widest point of ocular and approximately midway along its height, anterior margin barely covered by preocular plate in lateral view. Four postoculars bordering ocular and subocular between parietal and fourth supralabial. Three supralabials, third the largest, all except first with long axis oblique to long axis of body, first approximately square. Supralabial imbrication pattern T-V, posterior border of second supralabial overlaps anteroventral margin of preocular, that of third supralabial overlaps anteroventral margin of subocular. Mental hexagonal, wider than long, projecting slightly beyond curve of lower jaw and fitting into notch on upper lip when mouth is closed. Infralabials two on each side. Head glands no longer obvious due to bleached nature of specimen.
Longitudinal scale rows 24/22/20; mid-dorsal scale rows between rostral and tail tip 649; subcaudals 30; dorsocaudals 28; apical region with transverse, keratinized spine projecting posteroventrally at approximately 70˚ to axis of anteroventral surface of that terminal scale (and, hence, to body axis).
In preservative, dorsum very pale brown, bleached, with no difference between dorsal and ventral surfaces. Iris black; pupil pale gray, with black circle interior to that.
Variation. The sole additional specimen that is complete is YPM 9066 (AMNH 66739 comprises only the head and anterior portion of the body, cf. McDowell 1974). This specimen is smaller than the holotype but retains some vestiges of color pattern. Sex male; gular region visible from side; superior nasal suture inclined at approximately 10˚ from horizontal; ocular extends dorsally well medial to the preocular; approximately 25% of eye covered by preocular. Longitudinal scale rows 24/22/20; mid-dorsal scale rows 664; subcaudals 27; dorsocaudals 24. Head glands visible on rostral, nasals, and preoculars, but barely discernible, uncountable. Other features same as holotype. In preservative, pale brown, dorsum very slightly darker than ventrum, with color gradually decreasing ventrally; each scale slightly darker anteriorly. L = 188 mm, SVL = 182 mm, TL = 6 mm, HW = 2.7 mm, NW = 2.7 mm, SN = 1.9 mm, SW = 2.5 mm, PSN = 0.9 mm, RW = 1.45 mm, EW = 0.40 mm, W = 3.3 mm, VW = 2.8 mm, TW = 2.7 mm, L/W 57, M = 1.3 g (in preservative).
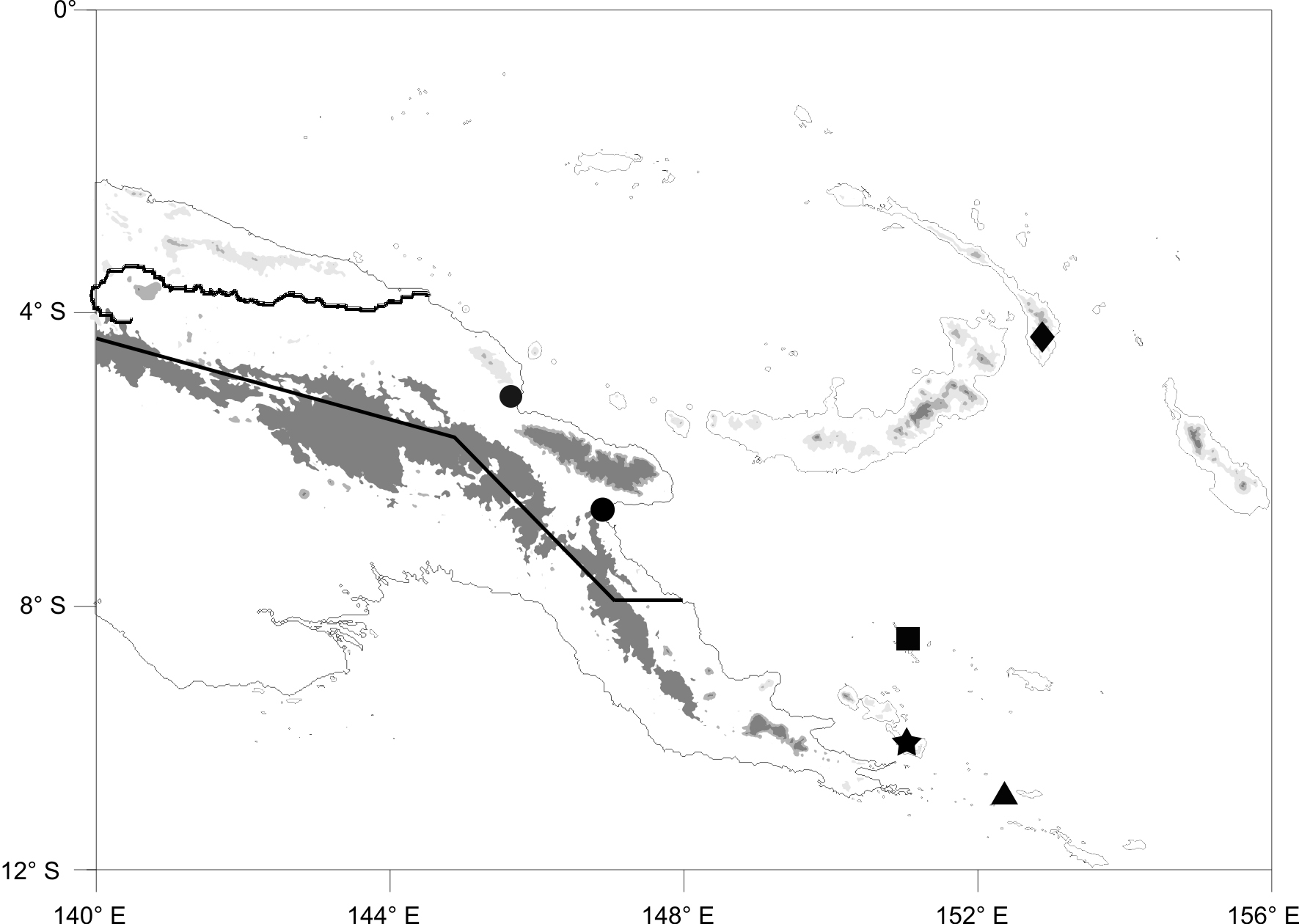
Distribution. The exact collection locality of the holotype of Typhlops depressiceps is unknown. Sternfeld (1913) merely noted that it came from “Neu Guinea,” which McDowell (1974) sensibly interpreted to mean German New Guinea, which was the northern portion of present-day Papua New Guinea ( Fig. 2 View FIGURE 2 ). The two specimens with known data come from Omuru (5.31˚S, 145.70˚E, YPM 9066), Madang Province, and Lae (6.73˚S, 146.99˚E, AMNH 66739), Morobe Province, Papua New Guinea ( Fig. 2 View FIGURE 2 ). The species is likely restricted to the northern and/or northeastern lowlands of New Guinea.
Remarks. McDowell (1974) included Typhlops monochrus Vogt 1932 in the synonymy of T. depressiceps . He did not examine the holotype, although he presumed it to reside in the ZMB collection. Vogt (1932) did not give an exact type locality for his species either, but noted that the vast majority of the collection he treated in the paper describing T. monochrus came from an expedition along the Sepik River ( Fig. 2 View FIGURE 2 ), and it has been reasonably presumed that the type specimen did as well ( McDowell 1974). The holotype has not subsequently been located in the ZMB ( Bauer et al. 2002), and it remains uncertain whether it still exists and, if so, where it resides. Consequently, it appears to have remained unexamined since Vogt’s original description, the brevity of which does not allow for useful taxonomic discrimination using modern diagnostic characters for the group.
McDowell’s (1974) synonymization of Typhlops monochrus with T. depressiceps may have been based on an error in Sternfeld’s (1913) original description of the latter, in which he claimed the holotype to have 24 rows of scales. Presumably this count was taken at midbody, although that is not clearly specified. Sternfeld’s report was repeated by de Rooij (1917) and McDowell (1974), neither of whom examined the type specimen. McDowell (1974) did remark that the single complete specimen of presumptive G. depressiceps that he did examine (YPM 9066) had 22 mid-body scale rows, which he thought served to connect Sternfeld’s T. depressiceps (24 scale rows) with Vogt’s T. monochrus (20 scale rows). It was partly this presumed annectance that led McDowell to synonymize the latter with the former. Based presumably on this history, Wallach (1996b) reported the number of longitudinal scale rows (LSR) in G. depressiceps to range from 20–24. He did not define LSR, but based on his reported values (and the contrast in values between what he reports for G. mcdowelli in his tables 1 and 2), it too evidently refers to a count taken at midbody, which is standard for snakes unless otherwise noted. This interpretation is consistent too with his use of mid-body scale counts in his dichotomous key provided in the same paper. In contrast to these reports, I find the holotype of T. depressiceps to have 22 scale rows at midbody, consistent with what McDowell and I have found for the Omuru specimen. Hence, there is no annectance in longitudinal scale rows between the assorted specimens I assign to G. depressiceps and the type description of Typhlops monochrus . Given the generally conserved nature of scale-row patterns among blindsnakes, there is, therefore, reason to entertain the notion that T. monochrus may prove to be a valid taxon differing from G. depressiceps in having 20 instead of 22 mid-body scale rows. Determining whether that possibility is valid will require collection of new material from northern Papua New Guinea inasmuch as the holotype remains unlocated and possibly lost. Nonetheless, future workers on these snakes should be aware that present evidence does not yet settle the status of T. monochrus .
An additional morphological discrepancy in the literature is that Wallach (1996b) reported the number of middorsal scale rows in Gerrhopilus depressiceps to vary from 631–713. Of the three specimens Wallach (1996b) examined, I obtain values of 627 (vs. his 631) for USNM 195953, 647 (vs. his 648) for MCZ 145954, and 649 (vs. his 713) for ZMB 23986 ( Table 1). Count differences for the first two specimens are trivial, but that for ZMB 23986 is significant. This large divergence between our observations for ZMB 23986 is important because Wallach’s value would imply a much smaller difference in mid-dorsal scale rows between G. depressiceps and two of the species described below than appears to be the case. I have repeated my counts for these specimens two or three times for each and I am confident that G. depressiceps , on the basis of available specimens, does not attain the large number of mid-dorsal scale rows attributed to it by Wallach (1996b). Consequently, I report a much narrower range in this character for G. depressiceps and the new taxa I remove from that species than appears from the data presented by him.
| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gerrhopilus depressiceps
| Kraus, Fred 2017 |
Typhlops monochrous
| Loveridge 1948: 320 |
Typhlops depressiceps
| Vogt 1932: 293 |
| Sternfeld 1913: 384 |