
Pseudopaludicola parnaiba, Roberto, Igor Joventino, Cardozo, Dario & Ávila, Robson W., 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3636.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:25E598D7-89E7-48DF-9F6E-32BBFD3DAA68 |
|
DOI |
https://doi.org/10.5281/zenodo.5665447 |
|
persistent identifier |
https://treatment.plazi.org/id/902A87C4-DA77-FFEC-FF54-F9BC8099FD21 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudopaludicola parnaiba |
| status |
sp. nov. |
Pseudopaludicola parnaiba , new species
Holotype (figs. 1–2). URCA 2160 (Adult male) collected on April 14 2012 by I.J. Roberto, R.W. Ávila, C.F. Da Silva and D.B. Oliveira, approximately 12 kilometers (in straight line) southeast of Ribeiro Gonçalves ( 07°35’42.15”S / 45°20’36.63”W; datum WGS 84), Municipality of Ribeiro Gonçalves, Piauí State, Brazil. Paratypes. Three adult females (URCA 2164, 2166, 2181), and 12 adult males (URCA 2158, 2162 3, 2165, 2168, 2174, 2178, 2180, 2182, 2184, 2191 2) collected on 14 April 2012 by the same collectors and at the same locality as the holotype, and eight adult males (URCA 2361 3; URCA 3864–8) collected on 18 January 2011 by I.J. Roberto, S.C. Ribeiro and J.A. Araújo Filho at the same locality as the holotype.
Diagnosis. Pseudopaludicola parnaiba sp. nov. is a member of Pseudopaludicola , based on the presence of one hypertrophied antebrachial tubercle on the external border of the forearms, posterolateral process of the hyoid outlined and epicoracoid cartilages slightly overlapped. The new species is characterized by the following: its small size (SVL in males 11.5–14.5, n =21; females: 13.0–14.3, n =3); body slender; toe tips knobbed with central groove; abdominal fold complete; tibio-tarsal articulation reaching the posterior border of the eye; prepollex and prehallux with base element and one segment; and advertisement call composed of a series of non-pulsed notes, with 6 to 46 notes per call and dominant frequency of 4794.7 ± 296.3 Hz.
Comparison with other species. The advertisement call with groups of 6–46 non-pulsed notes per call distinguishes P. parnaiba from all Pseudopaludicola species with pulsed advertisement calls: P. boliviana (Duré et al. 2004) , P. m u r u nd u (Toledo et al. 2010), P. s e r r a n a (Toledo, 2010) P. falcipes , P. mystacalis , P. saltica (Haddad and Cardoso, 1987) and P. m i n e i r a (Pereira & Nascimento, 2004). In relation to P. giarettai , the vocalization of P. parnaiba is distinguished by the presence of groups of 6–46 non-pulsed notes per call, while only one non-pulsed note per call (with four harmonics) is present in P. giarettai (Carvalho, 2012) . On the other hand, P. parnaiba dominant frequency of (4794.7 ± 296.3 Hz), and each note with an ascendant frequency at the beginning of the call, which gradually decreases to the end of the note, separates this taxon from P. hyleaustralis (dominant frequency 3938 ± 158.65 Hz, and each note with a slight ascendant frequency modulation in its first half, and another ascendant frequency modulation in its last half (Pansonato et al. 2012), and P. canga (dominant frequency 3692.1–4608.1 Hz, Giaretta & Kokubum, 2003; Pansonato et al. 2012) see Table 1 View TABLE 1 . The advertisements calls of Pseudopaludicola ceratophyes , P. l la ne r a, P. pusilla , P. riopiedadensis and P. ternetzi are undescribed.
Reference This study Pansonato et al. 2012 Pansonato et al. 2012 Carvalho, 2012 Additionally, the new species can be distinguished from all recognized Pseudopaludicola species by a combination of external morphology and osteological characters.
The smaller SVL of males ( 11.5–14.5 mm, n = 20) separates P. parnaiba from P. giarettai (range 16.2–18.0 mm, n = 10, Carvalho, 2012), P. canga ( 14.6–16.5 mm, n =12, Giaretta & Kokubum, 2003; Pansonato et al. 2012), P. ternetzi (16.0– 18.6 mm, n = 13, Lobo, 1996), P. saltica ( 15.2–16.9 mm, n = 9, Haddad & Cardoso, 1987), and P. serrana (15.0– 15.8 mm, n = 5, Toledo, 2010).
The slender body separates P. parnaiba from P. ternetzi , and P. riopiedadensis (which have a more robust body aspect; Lobo, 1995, 1996; Giaretta & Kokubum, 2003), and P. mineira (globular body aspect; Lobo, 1994, 1995).
The fingertips knobbed, distinguish P. parnaiba from the species assigned to the P. pusilla group ( P. b o l i v i a n a, P. ceratophyes , P. llanera , and P. pusilla ), which have T-shaped terminal phalanges (Lynch, 1989; Lobo, 1995, Cardozo & Suarez, 2012).
The presence of a complete abdominal fold differentiates Pseudopaludicola parnaiba from P. falcipes (incomplete or absent abdominal fold, Lobo, 1994, 1995).
The tibio-tarsal articulation reaching the posterior border of the eyes separates P. parnaiba from P. saltica , P. murundu and P. serrana , where the tibio-tarsal articulation extends beyond the tip of the snout (Lobo, 1994, 1995; Toledo, 2010; Toledo et al., 2010).
The prepollex composed of the base element and a long mineralized segment distinguishes P. parnaiba from P. falcipes , P. saltica , and P. t e r n e t z i (prepollex with base and two elements; Lobo, 1995). The prehallux formed by the base and one mineralized segment separates P. parnaiba from P. boliviana , P. llanera , P. mystacalis , P. pusilla , and P. t er ne tz i (prehallux with base and two mineralized segments; Lobo, 1995).
Although P. ameghini was referred to as valid species on the basis of karyological features by Fávero et al. (2011), there is no a formal revalidation of this taxon. According to Lobo (1996), the type specimens of P. ameghini and P. m y s t a c a l i s collected by Cope (1887) are of similar external morphology, and should be considered as synonyms. In this sense, a taxonomic revision of the types specimens housed at ANSP in the framework of the new evidence available (Duarte et al. 2010; Fávero et al. 2011) is necessary to elucidate the real taxonomic status of P. ameghini . For this reason this taxon is not included in the comparisons of P. parnaiba , as in other recent descriptions of Pseudopaludicola species (Pansonato et al, 2012, Carvalho, 2012).
On the other hand, although the advertisement calls of P. ternetzi and P. riopiedadensis are still undescribed, both taxa present pulsed advertisement calls according to Cardozo & Toledo (unpublished data). This character state and the external morphology (body size and slender aspect) presents in P. parnaiba are important characters to separate the new species from the taxon called P. riopedadensis
Description of holotype. Head longer than wide (cephalic index: head length/width = 1.24). Snout acuminate in dorsal view, protruding from the jaw in lateral view. Nostrils nearer to the tip of snout than to the eyes. Internarial distance smaller than interorbital space. Tympanum not visible. Superficial supratympanic fold. Undifferentiated canthus rostralis. Upper lip with white vertical stripes. Choanae rounded. Circular pupil. Tongue entire, oval, free behind without pigment at the base. Vocal sac single. Premaxillary and maxillary teeth present. Body slender. Skin smooth, with small flattened glandular warts on the flanks. An X-shaped glandular fold is evident in the interscapular region. Abdominal fold complete, thoracic fold present and gular fold indistinct. Antebrachial tubercle evident. Inner and outer metacarpal tubercles elongated and rounded, respectively. Length of fingers IV<II<I<III, with distal subarticular tubercles developed. Small supernumerary tubercles present at the base of the subarticular tubercles in the hand. Nuptial pads light cream, poorly developed, nuptial asperity present on the base of the thumb. Arms speckled dorsally, with dark crossbar. Inner metatarsal tubercles well developed, curved towards the tip of the leg, outer metatarsal tubercle conical and perpendicular to the plane of the foot. Tarsal fold slightly curved, with a thickening at the middle of the tarsus, simulating a tubercle. Toe webbing fringed. Length of the toes IV>III>V>II>I, with tips not expanded laterally.
Measurements of holotype (in mm). SVL 13.6; HW 4.6; HL 5.7; TL 7.0; THL 6.6; FL 8.1; ED 1.7; IOD 1.5; IND 1.1; NSD 0.9; END 1.2.
Color of the holotype. In life, dorsum brown, with two dorsolateral bands poorly defined, extending from postocular to inguinal regions. A thin white vertebral line extends from the tip of snout to cloaca. Belly white; dorsal view of arms uniform brown with two dark blotches; ventral region white, with scarce dark spots on chest, thighs and arms. The hind limbs have a dorsal striped pattern. The inner thighs have a light brown background finely spotted with black. Vocal sac yellowish with dark dots. Upper lip with white vertical stripes. Iris pale bronze with a black vertical line below the pupil. In preservative, dorsum and dorsal surface of limbs grayish brown, with dark dots on the ventral surface of legs and arms. Belly white; vocal sac white with scattered dark dots. The postocular gray stripe is less evident.
Variation. The dorsal color pattern varies from uniform brown to marble, with poorly defined dorsolateral bands. In most individuals of type series (87%), the vertebral line is absent. In some individuals without vertebral line (32%), a brown interorbital band is present. The paratypes (URCA 2162–4, 2166, 2174–6, 2178, 2180–2, and 2191) have a gray band perpendicular to the postocular region, reaching the upper arm. The presence/absence of Xshaped glandular fold varies among the paratypes. Eight individuals (URCA 2158, 2163–4, 2176, 2180, 2184, 2362–3) have a second antebrachial tubercle, always smaller than the first one and placed near the elbow. The paratype measurements as “mean ± standard deviation (range)” for 21 males were SVL 12.9 ± 0.9 (11.5–14.5), HL 4.7 ± 0.5 (4.0–5.6), HW 4.2 ± 0.4 (3.5–4.9), IND 1.1 ± 0.1 (0.9–1.2), NSD 0.7 ± 0.2 (0.4–1.0), END 1.1 ± 0.2 (0.9–1.4), ED 1.6 ± 0.1 (1.3–1.8), IOD 1.3 ± 0.1 (1.1–1.5), THL 5.7 ± 0.4 (5.0–6.5), TL 6.7 ± 0.4 (6.0–7.8) and FL 7.8 ±0.8 (7.1–10.2), and for three females SVL 13.6 ± 0.7 (13.0–14.3), HL 4.7 ± 0.3 (4.5–5.1), HW 4.5 ± 0.4 (4.1–4.8), IND 1.1 ± 0.1 (1.0–1.1), NSD 0.8 ± 0.1 (0.7–0.9), END 1.2 ± 0.3 (1.0–1.3), ED 1.7 ± 0.1 (1.6–1.8), IOD 1.5 ± 0.2 (1.3–1.6), THL 6.3 ± 0.5 (5.8–6.8), TL 7.2 ± 0.2 (7.0–7.3) and FL 8.2 ± 1.1 (7.0–9.1).
Osteology (fig. 3). Skull longer than wide. Complete maxillary arch. Alary processes of premaxillae directed dorsally, parallel to each other (frontal view). Premaxillae bearing 7–9 teeth and maxillae with 24–28 curved and pedicellated teeth. Nasal narrow and ovoid, with irregular inner margin and ending pointed posteriorly. The nasals are separated from each other and do not overlap the maxillary. In dorsal view, the nasal bone does not overlap the sphenethmoid. Anterior margin of the sphenethmoid not reaching the middle of the choana, posterior margin Ushaped. In dorsal view, sphenethmoid short with a thin internasal septum. Frontoparietals with regular inner margins that leaves the frontoparietal fontanelle partially exposed in the frontal region (frontoparietals with regular inner margins, leaving the frontoparietal fontanelle mostly hidden in URCA 3864). Optic foramen large sized, ovoid and located at the junction between the prootics and sphenethmoid. Oculomotor foramen of narrower diameter, located posterior to the optic foramen. Prootic foramen oval, placed posterior to the optic capsule. Jugular foramen composed of two holes of similar diameter. Triradiate pterygoid. The posterior ramus of the pterygoid is longer than middle and separated from the quadrate; medial ramus detached from the wings of the parasphenoid and supported by cartilage in the otic region of the skull. Anterior ramus separated from the palatine. Quadratojugal with anterior process reduced. Triradiate parasphenoid, with the tip of the cultriform process irregularly shaped. Parasphenoid wings directed backwards. Neopalatines poorly developed, investing the pars fascialis of maxillae. Prevomers irregularly shaped, wider in their middle region and without teeth. Squamosal with the ventral ramus wider at its ventral end, zygomatic ramus shorter than otic, with a descending process well developed. Tympanic annuli cartilaginous, opened dorsally. Plectrum of uniform width. Operculum cartilaginous, rounded. Mandible with a thin dentary covering the lateral margins of Meckel’s cartilage. The inner part of this cartilage is covered by the angular bone, which is thin and without coronoid process evident. Articular region of lower jaw cartilaginous, dentary plain, without denticle-like structures.
The larynx ( Fig. 3 View FIGURE 3 C) has the arytenoid as two small oblong shells and the cricoid forms a complete ring. Esophageal process short and thin; muscular and bronchial processes thin. Cardiac and articular processes poorly developed.
Hyoid plate cartilaginous, with dispersed mineralization ( Fig. 3 View FIGURE 3 C). Anteromedial processes of the hyoid short and divergent. Postero- and anterolateral processes vestigial and absent, respectively. Posteromedial processes with cartilaginous ends.
Vertebral column with eight procoelous, non-imbricate, presacral vertebrae ( Fig. 3 View FIGURE 3 D). Cotylar facets of the atlas separated ( type I of Lynch, 1971). Atlas body widest among the vertebrae. In dorsal view, the anterior margin of the atlas contacting presacral II through a wide neural process. Anterior portion of the illium articulating with the ventral surface of the sacral diapophysis. Pubis cartilaginous. Sacral diapophysis narrow and not expanded. Urostyle with dorsal spine well developed and exit of spinal nerve X not visible. Urostilar articulation bicondylar.
Pectoral girdle arciferal ( Fig. 3 View FIGURE 3 E), with the left epicoracoid cartilage slightly overlapping the right one, in ventral view (the right epicoracoid cartilage slightly overlaps the left in URCA 3868). Clavicle thin, posterior margin concave, separated from the acromial region of the scapula by an anterior projection of the procoracoid. Scapula with the pars acromialis and glenoidalis poorly developed. Anterior process of the suprascapula present. Omosternum cartilaginous, with expanded terminal end. Sternum simple, not bifurcated, with osseous mesosternum (mineralized in URCA 3865) and cartilaginous xiphisternum expanded.
Humerus with deltoid crest developed. Radius-ulna fused, leaving a narrow groove between them. Carpus composed of five elements: radial, ulnar, Y element, distal carpal V-IV-III and distal carpal II. Prepollex with osseous base and one mineralized long segment. Phalangeal formula: 2-2-3-3. Fingertips knobbed, with central groove.
Hind limbs with the tibio-fibula fused at the proximal ends. Tarsus composed of central, distal tarsal I, and distal tarsal II–III. Prehallux composed by an osseous base and one long mineralized segment. Phalangeal formula: 2-2-3-4-3. Toe tips knobbed with central groove ( Fig. 3 View FIGURE 3 F).
All specimens examined, have small osseous lateral projections at the anterior extremities of the metacarpal epiphysis and some phalanges. In the forelimbs, in dorsal view, there is a small rounded sesamoid bone over the distal carpal V-IV-III, embedded in the tendon of the muscle carpi ulnaris. In ventral view, the palmar sesamoid (in the m. digitorum longus) has an irregular quadrangular shape. Also are present glide sesamoids on the flexor tendons of the hands, at the junction of the proximal and medial phalanges in all fingers. The hind limbs have osseous lateral projections at the epiphysis of some metatarsals and phalanges, sesamoid graciella (at the union of femur with tibia-fibula, embedded in the tendon of the m. gracilis major); S. cartilage (in the tendon of the m. plantaris profundus) and two plantar sesamoids with ovoid shape and similar size, placed at the tendons that form the aponeurosis plantaris. Additionally, near the distal end of the sacral diapophyses, there are a couple of sesamoid bones (S. of the sacral vertebrae) with elongated shape and similar width as the diapophyses.
Advertisement call (fig. 4, Table 1 View TABLE 1 ). The individuals of P. parnaiba were recorded at night (18:00–20:00 h) on 18 January 2011 ( n =3) and 14 April 2012 ( n =2), with air temperature 26ºC. The advertisement call is composed of a series of non-pulsed notes (call series) with 11.7 ± 9.7 notes per call (6–46). The mean call series duration is 0.9 ± 0.9 sec (0.4– 3.9 s), emitted in short intervals from 0.4–1.8 sec (0.8 ± 0.5). The average number of notes per minute is 91.9 ± 41.9 (16–162). The mean note duration is 0.02 ± 0.02 sec (0.01–0.4) with inter-note interval of 0.07 sec ± 0.06 (0.04–0.6). The mean number of notes per second is 48.8 ± 12.6 (29.4–100). Each note has a slight ascendant frequency modulation at the beginning of the call, which gradually decreases to the end of the note. The species frequency range is 4338.5 ± 473.1 Hz (3295.7–4786.7) to 5364 ± 285.8 Hz (4788.5–6002.9), with dominant frequency of 4794.7 ± 296.3 (4220.5–5168).
Geographic distribution. Pseudopaludicola parnaiba is only known from its type locality, in the western region of Piauí State, bordering with Maranhão State, Northeast Brazil. This region is part of the Cerrado domain (sensu Ab’ Saber, 1977).
Natural history notes. Pseudopaludicola parnaiba was found in a disturbed area close to the Parnaíba River in the months of February and April. On both field trips, we found adult males in calling activity. Individuals vocalized during afternoon and at late night. As other species of the genus, the individuals of P. parnaiba vocalized hidden on grass stems on muddy soil in swamp areas (e.g., Haddad & Cardoso, 1987; Lobo, 1996; Carvalho, 2012). Scinax fuscomarginatus , S. nebulosus , Hypsiboas multifasciatus and H. punctatus were also found in calling activity in the same area.
Etymology. The specific epithet parnaiba refers to the Parnaíba River, an allusion to the habitat of the species, which occurs along the banks of this river.
Remarks. Historically, the Pseudopaludicola species occurring in Northeast Brazil have been poorly studied. Lobo (1995, 1996) assigned the specimens from Ceará, Paraíba, Pernambuco and Bahia states, in Northeast Brazil to P. mystacalis .
Later on, Rodrigues (2005) mentioned two species of Pseudopaludicola inhabiting the Caatinga biome in Northeast Brazil, P. falcipes and P. mystacalis , which were later called as Pseudopaludicola sp.1 and P. sp. 2 by Caramaschi (2008). In Piauí State, the taxonomic status of Pseudopaludicola populations remains unknown. Caramaschi and Jim (1983) mentioned the occurrence of Pseudopaludicola falcipes for Picos municipality, a typical Caatinga area, and Loebmann and Haddad (2010) assigned two indeterminate species to this biome (referring to them as P. sp (gr. falcipes ) and P. sp (gr. mystacalis ). However, the species groups referred by Loebmann and Haddad (2008) are not natural groups. The only group supported by an apomorphic character (end of the phalanges T-shaped) is the P. pusilla group (Lynch, 1989; Lobo, 1995).
On the other hand, Da Silva et al. (2007) assigned to P. mystacalis specimens from the municipality of Ilha Grande, along the coast of Piauí, and later, Loebmann and Mai (2008) sampled seven localities in the same area and cited the occurrence of an unidentified species, which they called P. gr. falcipes . In this sense, a major integrative taxonomic revision of Pseudopaludicola species inhabiting Northeast Brazil is needed to determine the true taxonomic status of multiple populations historically assigned to P. falcipes , P. m y s t a c a l i s and/or indeterminate specimens of the genus.
In conclusion, several names have been assigned to different Pseudopaludicola populations. However, the distribution of P. falcipes involve Southern Brazil through southeastern Paraguay and Uruguay to northeastern Argentina (Frost, 2011), and the presence of this taxon at northern Brazil is discarded by our revision, while the complete range of P. mystacalis is not fully understand, being necessary a complete revision of this taxon involving multiple lines of evidence like morphology, bioacoustics and genetic data.
It is important to remark that the specimens considered as P. mystacalis in the phylogenetic analysis of Lobo (1995) correspond to at least three species with different chromosome numbers: P. mystacalis , P. canga , and the populations from Bahia (see Giaretta & Kokubum, 2003; Duarte et al. 2010; Fávero et al. 2011), being necessary to recode the characters used by Lobo (1995), including all the species described up to the present, in order to obtain a more robust and inclusive phylogenetic hypothesis.
The area of occurrence of Pseudopaludicola parnaiba , within the Cerrado domain, and the higher level of deforestation for soy and cellulosic biofuel production, as well as dam constructions, is an increasing threat for the species. Despite the presence of a protected area in the region, Uruçuí-Una Ecological Station, we still need to determine if the species occurs in this area. In this sense the new species must be considered as poorly known. New inventories and ecological data surveys in the region are needed to determine the true conservation status of this species.
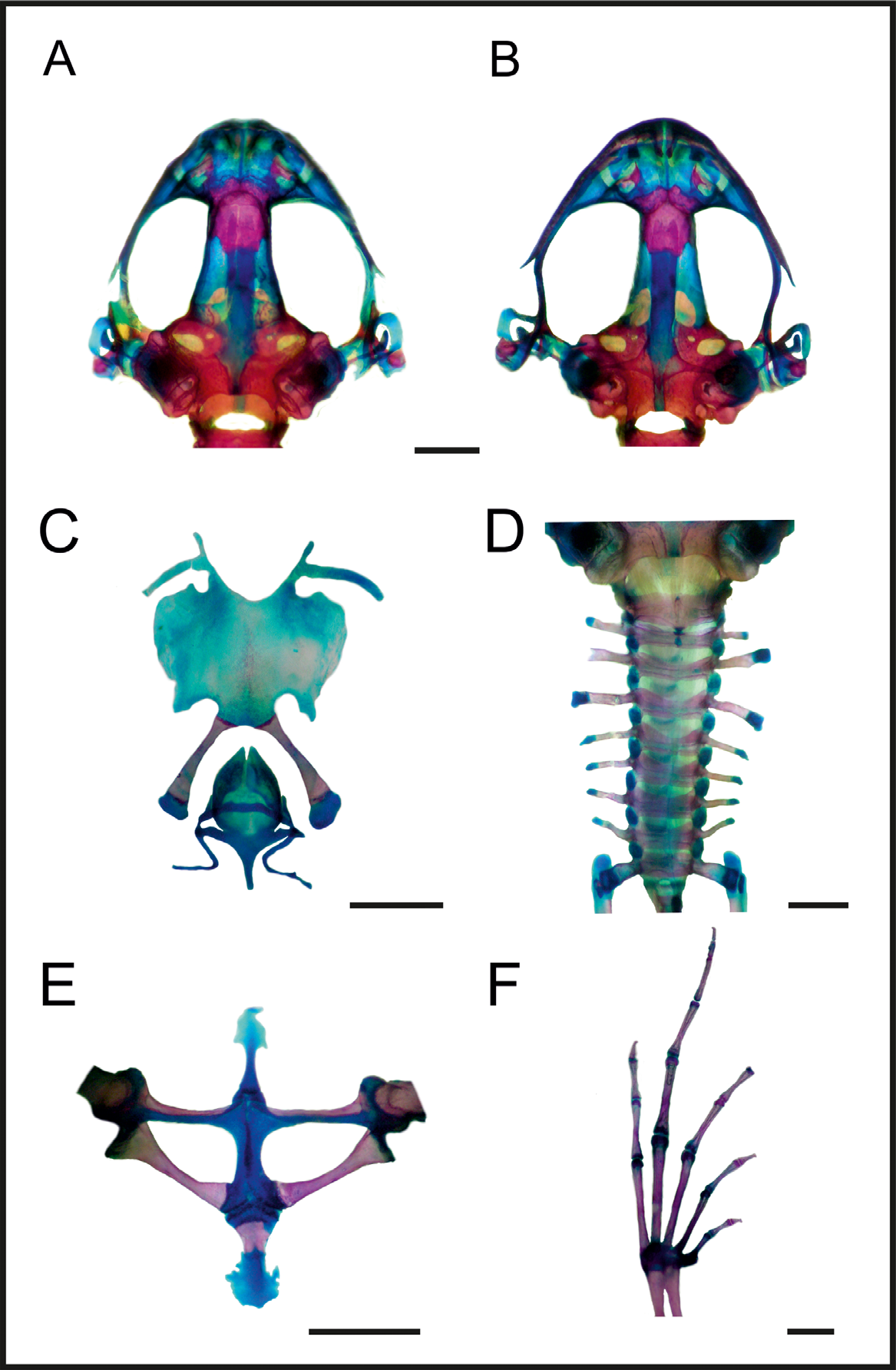
TABLE 1. Acoustic parameters of Pseudopaludicola species with non-pulsed advertisement call.
| Acoustic Parameters | Pseudopaludicola parnaiba (sp nov) | Pseudopaludicola hyleaustralis | Pseudopaludicola canga | Pseudopaludicola giarettai |
|---|---|---|---|---|
| Call duration (s) | 0.9 ± 0.9 (0.4–3.9) | 4.25 ± 2.1 (1.1–8.2) | 0.2–1.3 | 0.15 + 0.016 (0.12–0.19) |
| Note duration (s) | 0.02 ± 0.02 (0.01–0.4) | 0.03 ± 0.003 (0.025–0.05) | 0.014–0.03 | * |
| Inter note duration (s) | 0.07 ± 0.06 (0.04–0.6) | 0.08 ± 0.02 (0.035–0.2) | 0.04–0.11 | * |
| Intercall intervall (s) | 0.8 ± 0.5 (0.4–1.8) | 46.8 ± 10.4 (29.7–58.8) | 0.25–6.7 | 0.3 + 0.05 (0.18–1,3) |
| Note rate (min) | 2762 ± 503 (1764–6000) | 561.4 ± 41.9 (503.8–623.2) | 689.6–944 | 136.2 + 14.8 (117–153) |
| Dominant Frequency (Hz) | 4794.7 ± 296.3 (4220.5–5168) | 3938.9 ± 158.6 (3605–4164.3) | 3962.1–4608.1 | 4374 + 150.0 (3981–4719) |
| Minimum Frequency (Hz) | 3295.7 | 3562.3 | 3297 | * |
| Maximum Frequency (Hz) | 6002.9 | 4379.8 | 5238 | * |
| Air Temperature °C | 26 | 24 | 25–29 | * |
| Locality | Ribeiro Gonçalves, State of Piauí | Alta Floresta, State of Mato Grosso | Marabá, Conceição do Araguaia e Curionópolis, State of Pará | Curvelo, State of Minas Gerais |
| Country | Brazil | Brazil | Brazil | Brazil |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |