
Pliobates cataloniae, David M. Alba, Sergio Almécija, Daniel DeMiguel, Josep Fortuny, Miriam Pérez de los Ríos, Marta Pina, Josep M. Robles & Salvador Moyà-Solà, 2015
|
publication ID |
https://doi.org/ 10.1126/science.aab2625 |
|
publication LSID |
lsid:zoobank.org:pub:4A5EC1F1-29BD-4925-8CC6-04AB083C61DA |
|
DOI |
https://doi.org/10.5281/zenodo.4420189 |
|
persistent identifier |
https://treatment.plazi.org/id/3FB80ECA-174A-4D5E-9699-E5C960F2A17B |
|
taxon LSID |
lsid:zoobank.org:act:3FB80ECA-174A-4D5E-9699-E5C960F2A17B |
|
treatment provided by |
Plazi |
|
scientific name |
Pliobates cataloniae |
| status |
gen. et sp. nov. |
Pliobates cataloniae gen. et sp. nov.
Holotype: IPS58443, a partial skeleton with an associated skull ( Fig. 1 View Fig. 1 and movie S1), housed at the ICP. It is composed of 70 bones and bone fragments (table S1) found in close spatial association, which, given the lack of repeated elements, are attributed to a single adult female individual (based on the small canine alveolus), with an estimated body mass of 4 to 5 kg (tables S7 and S8). It includes large portions of the cranium with postcanine maxillary teeth ( Table 1), a mandibular fragment, a partial left forelimb (nearly complete humerus, radius, partial ulna, carpals, and bones of the manual rays), more fragmentary elements of the right forelimb, and bones from the hind limb.
Type locality: ACM/C8-A4 (els Hostalets de Pierola, Catalonia, Spain), in the ACM stratigraphic series (Vallès-Penedès Basin, northeast Iberian Peninsula).
Age, stratigraphic position, and distribution: Only known from the type locality, which has an estimated age of 11.6 Ma (middle/late Miocene boundary) and is thus somewhat younger than all other ACM hominoid- and pliopithecoid-bearing localities (9, 31, 33, 77, 78), the latter of which have been dated to 11.7 to 11.9 Ma [updated from (77, 78)].
Etymology: Genus name from the Latin plio- (itself from the Greek, meaning “greater in extent”) and from the Greek bates (meaning “the one that walks or haunts”). The name is a contraction of the genus names Pliopithecus (“more ape”) and Hylobates (“the one that walks in the woods or in the trees”), in allusion to the small body size and the mosaic of primitive (stem catarrhine–like) and derived (crown hominoid) features displayed by the new taxon. The species epithet is the genitive of the female substantive “Catalonia,” the Latin name of Catalunya (in which the type locality is situated).
Diagnosis
Small-bodied catarrhine primate (estimated female BM of 4 to 5 kg). Dental formula 2.1.2.3.
Female upper canines moderately compressed. Upper cheek teeth low-crowned and with subpyramidal, moderately peripheral, and inflated cusps. Upper premolars relatively broad and ovoid, P4 smaller than P3, both with heteromorphic cusps, a markedly convex lingual contour and a distinct lingual cingulum (more developed in the P4), a distinct transverse crest separating the restricted mesial fovea from the extensive trigon basin, and the postparacrista forming an abrupt angle with the distal marginal ridge. Upper molars only moderately broader than long, with markedly convex lingual profiles; buccal cusps quite peripheral and buccal cingula discontinuous; lingual cingula relatively well developed, shelf-like, and C-shaped, but not surrounding the hypocone (which is distinct and more peripheral than the protocone); mesial fovea restricted, with an obliquely directed preprotocrista, and trigon basin extensive, being separated by a continuous crista obliqua from the slightly smaller distal fovea, which displays no hypocone-metacone crest. M2 slightly larger than the M1, and M3 shorter and trapezoidal (due to the oblique buccal margin, with a centrally situated metacone and a rudimentary hypocone).
Face small but with a distinct snout, the anterior portion of the nasals being almost parallel to the palate. Maxillary sinus large and frontal sinus present but small. Nasal aperture narrow. Nasoalveolar clivus short, with an open palatine fenestra. Anteriorly slightly narrow palate with somewhat convergent upper tooth rows. Zygomatic root moderately high. Orbits subcircular, large, and frontated, with telescopic orbital rims located over the P4. Estimated cranial capacity (69 to 75 cm3) indicating a monkey-like degree of encephalization. External auditory meatus tubular but short and not completely ossified, with a V-shaped end and its anterior portion fused with the postglenoid process. Carotid foramen perforating the bulla posterodistally, and carotid canal horizontally and anteriorly oriented. Spinosum and postglenoid foramina absent. Jugular foramen large and ventrally visible.
Humerus without entepicondylar foramen and capitular tail, with a well-developed capitulum, and a narrow and deep zona conoidea.Radial head rounded and not very tilted, with a markedly beveled surface for articulation with the humeral zona conoidea, the articular surface for the ulnar radial notch extending along a large portion of the radial head, and a laterally facing bicipital tuberosity. Distal radioulnar joint fully diarthrodial, with an expanded and two-faceted semilunar articulation on the ulnar head, and a partially developed ulnar fovea. Ulnar styloid process with reduced girth and not articulating with the short pisiform. Triquetrum small and with a reduced articular surface for the ulnar styloid process. Hamate relatively long proximodistally, with a steep triquetrum facet, a relatively large head and a distally projecting hamulus. Capitate with a relatively small and oblong head and a divided facet for the second metacarpal on its radial side.
Differential diagnosis
The new taxon differs from pliopithecoids and dendropithecids in its lack of a humeral capitular tail, its hominoid-like proximal radial morphology, its expanded ulnar head with a two-faceted semilunar articulation, and its partially developed ulnar fovea. It further differs from these taxa and proconsulids in its more hominoid-like carpal morphology (including the lack of a pisiform facet for the styloid process, a capitate facet for the second metacarpal divided by a deep ligamentary notch, and a distally projecting hamulus in the hamate), and particularly from pliopithecoids in its overall larger muzzle, more horizontal nasals anteriorly, some details of the upper molars, and (at least compared with Epipliopithecus) the lack of an entepicondylar foramen in the humerus. It also differs from all of the above-mentioned taxa in its fused ectotympanic and postglenoid process, and from these taxa and hominids in its horizontal and anteriorly oriented carotid canal. Last, it differs from crown hominoids (hylobatids and hominids) in its incompletely ossified ectotympanic and in its more primitive dentition and forelimb morphology (particularly in the humeroulnar articulation).
Description, comparisons, and paleobiology Dental morphology and diet
Although the lack of lower dentition precludes comparisons with some taxa, the upper cheek teeth of Pliobates ( Fig. 1 View Fig. 1 D and Table 1) generally resemble those of other small-bodied Miocene catarrhines in both occlusal morphology and proportions (figs. S1 and S2). In contrast, they display a more primitive morphology than those of extant hominoids, including the similarly sized gibbons. Hylobatids possess more elongated cheek teeth with more peripheralized cusps, less developed cingula, and a much more extensive central fovea. Compared with Miocene small-bodied catarrhines from Eurasia and Africa (fig. S1), the upper molars of Pliobates more closely resemble those of the dendropithecid Micropithecus (21, 34, 35, 79) in several features, such as the markedly convex lingual profiles and moderately developed buccal cingula (albeit to a lesser extent than in Micropithecus), the C-shaped lingual cingulum that is mostly restricted to the protocone (not surrounding the hypocone), the well-developed and lingually situated hypocone, and the relatively narrow M1 and M2. Nevertheless, Pliobates differs in several features from Micropithecus, which has more restricted buccal cingula, a hypocone-metacone crest, and a relatively longer and less trapezoidal M3. The dentition of Pliobates more clearly differs from Epipliopithecus and other pliopithecoids (fig. S1), including from Barberapithecus [also recorded at the Vallès-Penedès Basin (36)] and Pliopithecus [previously recorded at ACM (33)] in several features, such as the more convex lingual profile, the more peripheral buccal cusps, the less developed cingula of the molars, the narrower M1 and M2, and the M3 occlusal morphology and proportions (fig. S2).
With regard to microwear features, the M1 displays a pitting percentage of 30.0%, a pit breadth of 5.67 μm, and a striation breadth of 1.98 μm. Based on pitting incidence ( Fig. 2 View Fig. 2 , A and B), which is the most useful metric for distinguishing among dietary categories (70), Pliobates closely resembles extant frugivores ( Pan troglodytes ) and eclectic feeders ( Papio cynocephalus ) that largely rely on ripe fruit. In contrast, the pitting incidence of Pliobates is higher than in extant folivores and much lower than in extant hard-object feeders (including orangutans). Compared with other extinct catarrhines from Western Europe, the pitting percentage of Pliobates is somewhat lower than in most pliopithecoids and hominoids, for which some degree of sclerocarpy has been inferred (66, 68). This low pitting incidence is consistent with the pit- and striation-breadth measurements (which show no sign of extreme folivory or specialized hard-object feeding) and most closely approaches that of the fossil hominoids Anoiapithecus brevirostris and Hispanopithecus laietanus, previously interpreted as soft frugivores (68). These results are confirmed by a multivariate analysis that simultaneously examined the three microwear variables ( Fig. 2 View Fig. 2 C and tables S9 to S11), in which Pliobates falls closer to the extant frugivorous–mixed-feeder centroid for the first and second canonical axes and is classified as a frugivore. Dental microwear analyses therefore indicate a mainly frugivorous diet for Pliobates , compatible with a high consumption of ripe fruit and a low sclerocarpic component.
Body mass
Dental BM estimates for the female holotype of Pliobates (table S7) range from 2.9 to 4.8 kg, with an average BM estimate of 3.9 kg and an uncertainty degree (based on the combined 95% confidence intervals for each dental locus) of 2.5 to 5.7 kg. Postcranial BM estimates (table S8) are on average 4.8 kg (range: 4.0 to 5.6 kg), based on catarrhine regressions, and 4.3 kg (range: 2.6 to 6.3 kg), based on hominoid regressions (estimates for each postcranial estimator and their confidence intervals are given in table S8). Given that the size of Pliobates is in the lower range for extant hominoids, the catarrhine regressions probably yield more accurate estimates, although the hominoid-based estimates are closer to the dental ones. Overall, the body mass of the holotype of Pliobates cataloniae can be estimated at ~4 to 5 kg (much lower than that estimated for Epipliopithecus vindobonensis , ~11 to 12 kg; table S8).
Cranial morphology and encephalization
A 3D virtual reconstruction of the cranium, based on the preserved specimens, is shown in fig. S3, whereas the final reconstruction (including mirrored portions) is shown in Fig. 3 View Fig. 3 and movie S1. Based on this reconstruction, the cranium of Pliobates differs from the primitive catarrhine condition (22, 37, 60) by being short, wide, and high. However, the tubular ectotympanic is short and incompletely ossified—i.e., less developed than in Saadanius and extant crown catarrhines (20–22). The maxillary sinus is extensive, as in stem catarrhines and hominoids (22, 80), and there is also a small frontal sinus, as in stem hominoids but unlike in stem catarrhines, cercopithecoids, hylobatids, and pongines (22, 37, 80). The face is short and displays anteriorly situated orbits, as in hylobatids, colobines, and some extinct small-bodied catarrhines such as Epipliopithecus, Micropithecus, and Lomorupithecus (19, 21, 38, 76, 79). However, Pliobates differs from these taxa (and more closely resembles hylobatids) by displaying a more well-defined muzzle (especially compared with Epipliopithecus) with long and more horizontal nasals, a higher zygomatic root (moderately high as in hylobatids, but less so than in hominids), an interorbital pillar nearly orthogonal to the frontal squama (as in hylobatids and chimpanzees), a high degree of orbital convergence and frontation (as in all extant hominoids), and thin and anteriorly projecting (telescopic) orbital rims [to a greater extent than in Epipliopithecus (38), and thus most closely resembling hylobatids and, as far as it can be ascertained with incomplete preservation, Micropithecus (79)]. Pliobates also displays derived hominoid features in the basicranium ( Fig. 4 View Fig. 4 A), including the absence of a postglenoid foramen with a large and ventrally visible jugular foramen (as in all extant hominoids), the foramen ovale situated anteriorly and laterally to the Eusta- chian aperture (as in hylobatids and African and African apes), and the horizontal and anapes), the fusion between the auditory meatus teriorly directed carotid canal in the petrosal bone and the postglenoid process (as in hylobatids (as in hylobatids).
Braincase measurements yield an average cranial capacity estimate of 69.0 cm3 (range: 41.4 to 110.7 cm 3; table S12), which is close to the estimates of 60.1 and 65.3 cm3 delivered by the two most reliable estimators (57) and only slightly lower than the estimate of 75.1 cm3 obtained from foramen magnum area (table S12). According to our estimates of body mass (4.5 kg) and cranial capacity [72 cm3 (average of cranial and foramen magnum estimates)], Pliobates would display a monkey-like degree of encephalization extensively overlapping with extant cercopithecoids (fig. S4 and table S13), being much more encephalized than the stem catarrhine Aegyptopithecus, slightly more so than the stem cercopithecoid Victoriapithecus, and only slightly less so than hylobatids and the extinct hominoids Proconsul and Oreopithecus. All these taxa, like cercopithecoids, are less encephalized than the extinct hominoid Hispanopithecus (Rudapithecus) and the extant great apes. Although humans are outliers in brain size–body size allometric regressions, great apes further display an allometric grade shift compared with hylobatids (and Pliobates ), which are only slightly more encephalized on average than cercopithecoids (58).
Postcranial morphology and locomotion
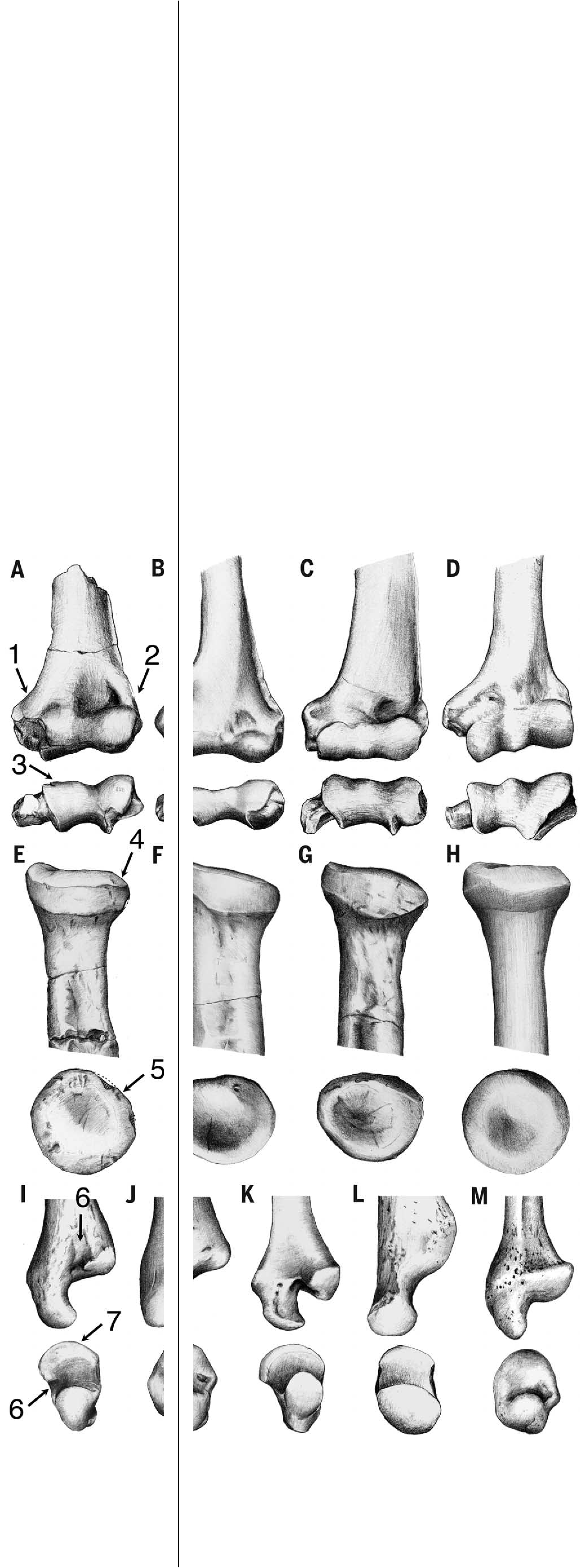
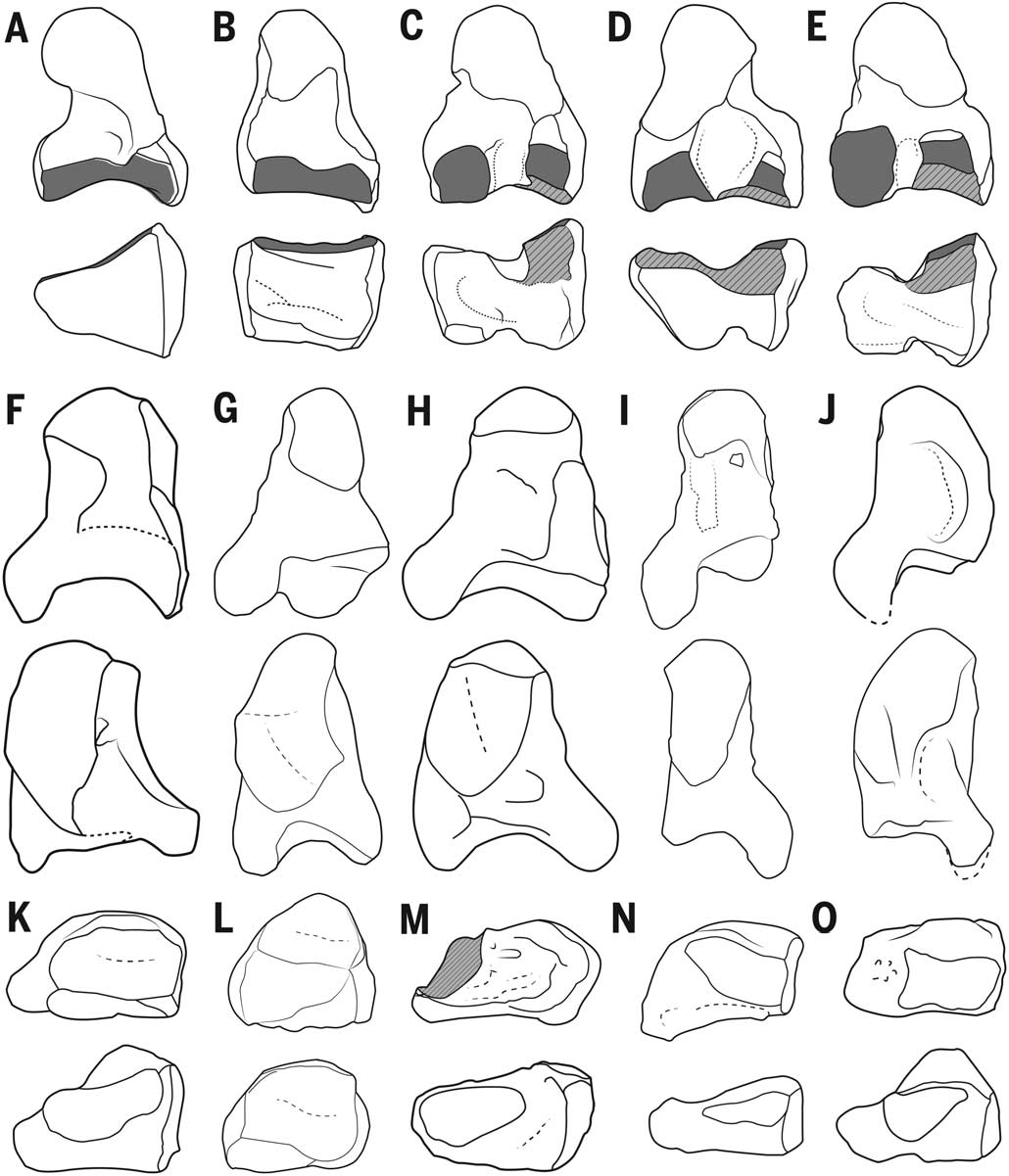
The humerus ( Fig. 5 View Fig. 5 ) resembles that of extant crown catarrhines, proconsulids, and dendropithecids by lacking (unlike Epipliopithecus ) an entepicondylar foramen (17, 20, 21, 38, 79, 81, 82). Pliobates more closely resembles extant hominoids in the laterally facing bicipital tuberosity in the radius (81, 83) ( Fig. 5 View Fig. 5 ), as well as in the configuration of the humeroradial articulation (81, 82, 84) ( Fig. 6 View Fig. 6 ), including: in the humerus, the lack of capitular tail [present in Epipliopithecus , dendropithecids, and cercopithecoids (84)] and the moderately globulous (although not posterolaterally expanded) capitulum with a well-developed zona conoidea [lacking in Epipliopithecus and dendropithecids (81–84)]; and, in the radius, the only slightly tilted and almost circular radial head with a small and flat area, a reduced lateral lip, and a beveled surface for the humeral zona conoidea. Pliobates also has a hominoid-like diarthrodial distal radioulnar joint (85–87), with a two-faceted expanded semilunar articulation in the ulnar head ( Fig.6 View Fig. 6 ). In this regard, Pliobates departs from Epipliopithecus, dendropithecids, and cercopithecoids (17, 38, 82) and more closely resembles Proconsul (88), although the ulnar head is less extensive than in extant hominoids. In contrast with these derived features, the humeral shaft and humeroulnar joint are plesiomorphic: The former ( Fig. 5 View Fig. 5 ) is anteriorly straight and somewhat proximally retroflexed; the latter ( Fig. 6 View Fig. 6 ) lacks the stabilizing features of extant hominoids (83, 89), as shown by the narrow ulnar trochlear notch without a median keel (in agreement with the absence of spooling and the poorly defined trochlear lateral keel in the humerus of Pliobates ).
Humeral torsion in Pliobates is estimated at 101°, irrespective of the method employed (based on the posterior buttress for the humeral head or the bisector of the bicipital groove), with a confidence interval spanning 95.7° to 106.3° [based on the prediction error (5.23%) for the bicipital groove method (50)]. This degree of torsion is moderate, higher than that estimated for Proconsul heseloni (92°), but comparable to estimates for Dryopithecus fontani (102°) and Dendropithecus macinnesi (103.5°), and only slightly below the value estimated for Epipliopithecus vindobonensis (109°). The humeral torsion of Pliobates is thus most comparable to that of non-atelid platyrrhines and lower than that of Ateles and extant hominoids, especially African great apes and humans [although the high degree of humeral torsion of extant hominoids is related to increased mobility at the glenohumeral joint, the higher values of great apes and humans appear related to knucklewalking and enhanced manipulation, respectively, rather than suspensory behaviors (50)]. In contrast to the moderate humeral torsion, the forelimb of Pliobates appears somewhat elongated relative to its body size (fig. S5). Allometric computations of relative forelimb length in fossils (residuals are given in table S14) must be considered with caution, because they are dependent on the accuracy of body-size estimates. However, the forelimb of Pliobates (based on our BM estimate of 4.5 kg) appears more elongated than that of Epipliopithecus (based on our estimate of 11.5 kg). The latter taxon, contrary to previous assertions (81, 83), has the generalized proportions of quadrupedal monkeys. Pliobates , in contrast, has a forelimb elongation similar to that of female orangutans and Brachyteles , although it is less extreme than in Ateles and especially than in hylobatids. The same pattern holds when the humerus and radius are analyzed separately, although in Pliobates , relative length is somewhat higher for the radius than for the humerus. Pliobates further displays a high arm angle (8°), which is considerably greater than the average in most anthropoids, except Hylobates (9.8°), Pongo (6.3°), and Ateles (6.5°) (51).
The ulnocarpal articulation of Pliobates is completely different from that of Epipliopithecus and dendropithecids (17, 38, 90), including a partially developed ulnar fovea ( Fig. 6 View Fig. 6 ), which, in extant hominoids, is the attachment area of the triangular disc ligament and the intra-articular meniscus (85–87). The ulnar styloid process is relatively long and slender, with no discernible articular surfaces for the pisiform or triquetrum. This agrees with the lack of an articular facet for the styloid process on the pisiform, like extant hominoids but unlike monkeys and Proconsul (91). However, in contrast to Pierolapithecus (92), the triquetrum of Pliobates shows a proximal articular facet, which is more developed than that present in hylobatids and sometimes Pan (51, 87) but less developed than in monkeys. This suggests that ulnotriquetral contact might have been reduced by some kind of intra-articular tissue, similarly to some Ateles species (93). Moreover, as in apes, the triquetrum of Pliobates is small relative to hamate size (fig. S6), indicating a reduced loading on the ulnar side of the wrist. However, as in monkeys and Proconsul, the triquetrum of Pliobates differs from that of extant apes and Pierolapithecus (92) by possessing a proximally protruding beak-like process ( Fig. 7 View Fig. 7 ). The hamate of Pliobates is “Miocene ape– like” (91, 92), although it more closely resembles that of hylobatids by possessing a dorsopalmarly narrow and proximodistally long triquetral articular surface that is proximally globular, as well as a distally projecting hamulus ( Fig. 7 View Fig. 7 ). Pliobates further resembles hylobatids and Ateles by having an oblong and mediolaterally narrow capitate head that, like the facet for the hamate, is proximodistally aligned. This morphology contrasts with the more globulous, wider, and ulnarly inclined capitate head of other catarrhines, including Proconsul (88, 91) and Pierolapithecus (92); in Pliobates , though, it is not radially inclined, as it is in hylobatids. Moreover, the capitate facet for the second metacarpal is divided by a deep ligamentary notch ( Fig. 7 View Fig. 7 ), as in extant hominoids and Pierolapithecus (92) but not in other catarrhines (including Proconsul), in which the facet for the second metacarpal is dorsopalmarly continuous and occupies the whole lateral aspect of the capitate ( Fig. 7 View Fig. 7 ) (91). Pliobates has a complex articulation between the third metacarpal and capitate, as in extant apes but not in Proconsul (91); however, as in Pierolapithecus (92), the capitate of Pliobates lacks a hook-like process.
Regarding positional behaviors, although the primitive morphology of the proximal humerus of Pliobates is suggestive of generalized abovebranch quadrupedalism (94), its overall postcranial body plan is more compatible with a locomotor repertoire that includes a large amount of cautious and eclectic climbing (87, 95). This inference is supported by the emphasis on pronation and supination capabilities, the reduced compressive forces transferred across the ulnar side of the wrist, and the important ulnar deviation and rotatory capabilities. It agrees with previous hypotheses on the original locomotor adaptations of hominoids (95, 96) and with recent interpretations of Proconsul that similarly depict this taxon as an arboreal quadruped with adaptations for cautious climbing and clambering (12, 88), including an incipient distal radioulnar diarthrosis that (unlike in Pliobates ) is still associated to a nonretreated ulnar styloid process (88). The reduced ulnocarpal articulation of Pliobates thus more closely foreshadows the condition of extant hominoids, although to a lesser extent than in the stem great ape Pierolapithecus (92), indicating a decreased emphasis on forearm use under weight-bearing conditions relative to Proconsul (88). Several characteristics of Pliobates (particularly the elongated forearm, the high arm angle, and the laterally facing bicipital tuberosity) further suggest some degree of below-branch forelimbdominated suspensory behaviors (51). However, the lack of hominoid-like elbow-stabilizing features in the humeroulnar joint (83, 89), the generalized metacarpophalangeal proportions, and the lack of marked phalangeal curvature suggest that Pliobates was not specifically adapted to perform the acrobatic suspensory behaviors (ricochetal brachiation) displayed by extant gibbons.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.