
Eremias dzungarica, Orlova & Poyarkov & Chirikova & Nazarov & Munkhbaatar & Munkhbayar & Terbish, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4282.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:7BBDADA0-FB34-4659-A97F-34C00583765E |
|
DOI |
https://doi.org/10.5281/zenodo.6029503 |
|
persistent identifier |
https://treatment.plazi.org/id/9316F96C-FFD8-873B-78D6-FDAD3B370DC5 |
|
treatment provided by |
Plazi |
|
scientific name |
Eremias dzungarica |
| status |
sp. nov. |
Eremias dzungarica sp. nov.
( Figs. 3–9 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ; Table 3)
Chresonymy:
“ Eremias multiocellata ” (partim) — Ananjeva 1972; Bannikov et al. 1977; Prokopov 1978; Brushko & Kubykin 1988; Brushko 1995; Orlova 1995; Orlova & Terbish 1997; Sindaco & Jeremcenko 2008; Tuniyev & Ananjeva 2010. “ Eremias multiocellata yarkandensis ” — Szczerbak 1974 (partim).
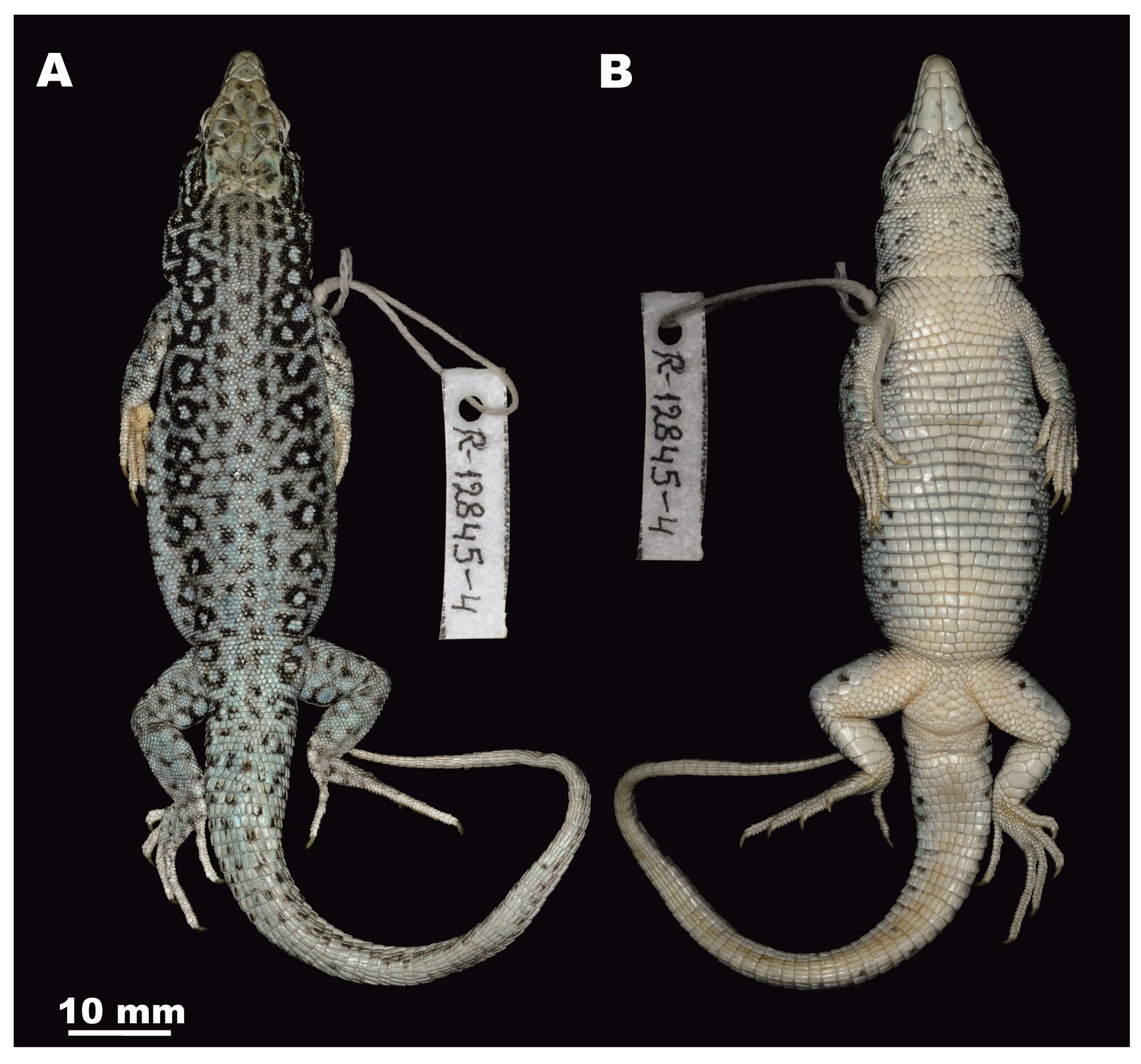
Holotype. ZMMU R-12845, adult male, individual lable R-12845-4; collected on 12.08.2008 from Mongolia, Khovd Aimaq, Khoovor, 7 km west from Uyench Sum (= Uench-somon ) (46° 05' N; 91° 56' E) by V.F. Orlova ( Figs. 3–6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). GoogleMaps
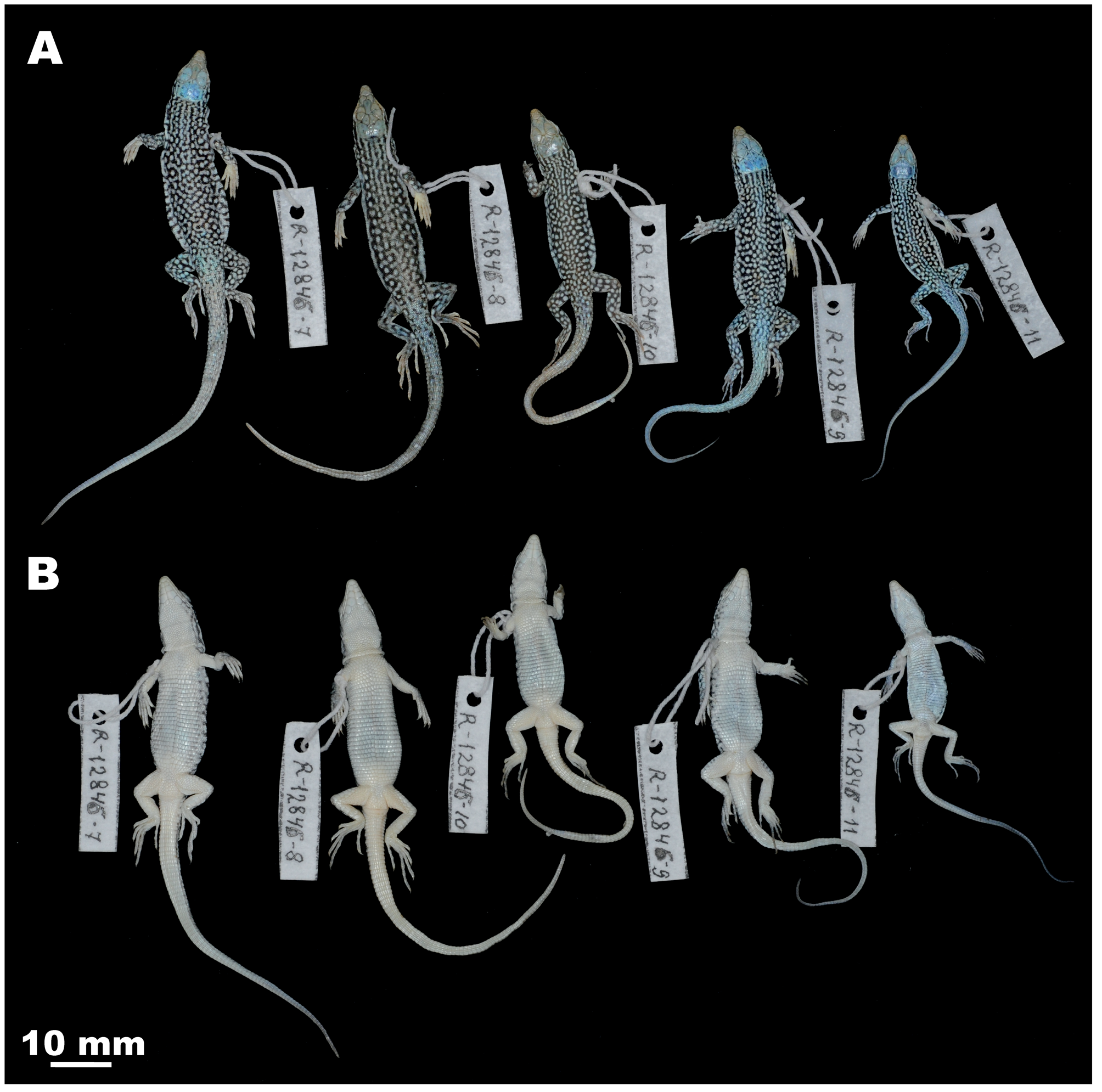
Paratypes. ZMMU R-12846-2 and R-12846-5, two adult males, and R-12846-6 one adult female; collection data same as for the holotype: Mongolia, Khovd Aimaq, Khoovor, 7 km west Uench-somon 46° 05' N; 91° 56' E; collected on 12.08.2008 by V.F. Orlova ( Fig. 7 View FIGURE 7 ) GoogleMaps . ZMMU R-12846-7–R-12846-11, 5 juveniles; collection data same as for the holotype: Mongolia, Khovd Aimaq, Khoovor, 7 km west Uench-somon 46° 05' N; 91° 56' E; collected on 12.08.2008 by V.F. Orlova ( Fig. 8 View FIGURE 8 ) GoogleMaps . ZMMU R-12862-1 and R-12862-3, two adult males, and R-12862-2, one adult female; collected from: Mongolia, Khovd Aimaq, Bulgan-Somon, urochishe Bayan-Mod, 11 km west Ikher-Toli , on 13.08.2008 by G. Davaasuren ( Fig. 7 View FIGURE 7 ).
Referred Materials. Mongolia: ZMMU R-5415 (1 male, 6 females) from Mongolia, Khovd Aimaq, 6-7 km to the West of Uyench Sum [ Uench-somon ]), coll. 05.07.1984 ; ZMMU R-5414 (1 male, 1 female) from the same locality as above, coll. 0 4.07.1984 (samples used in the allozyme study of Truweller et al. 1994) ; ZMMU R-5413 (1 male, 1 female), from the same locality as above, coll. 03.07.1984 ; ZMMU R- 5417 (2 females, 1 male), from the same locality as above, coll. 05– 06.07.1984 ; ZMMU R-5617 (2 adults), from the same locality as above, coll. 05- 06.07.1984 ; ZMMU R-6415 (1 male, 3 females), from Mongolia, Khovd Aimaq, env. Uyench Sum [ Uenchsomon ], 07.06.1984 ; ZMMU R-6417 (5 adults, 2 subad.), from Mongolia, Khovd Aimaq, 15 km E from Uyench Sum [ Uench-somon ] (Khuv-Us), coll. 07.06.1984 ; ZMMU R-6776 (2 females), from Mongolia, Khovd Aimaq, 15 km N from Uyench Sum [ Uench-somon ], coll. 10.08.1986 ; ZMMU R-5781 (2 males, 3 females), 6 km W of Uyench Sum [ Uench-somon ], coll. 23.07.1986 ; ZMMU R-5807 (1 male) from Mongolia, Khovd Aimaq, env. Uyench Sum [ Uench-somon ], coll. 23.07.1986 ; ZMMU R-8252 (4 males, 1 female) from Mongolia, Khovd Aimaq, 6-7 km to W from Uyench Sum [ Uench-somon ], coll. 04– 05.07.1984 ; ZMMU R-7617 (2 males), from the same locality as above, coll. 04.- 05.07.1984 ; ZMMU R-8449 (2 juv.) and ZMMU R-12550 (1 subad.) from Mongolia, Khovd Aimaq, 24 km N of Uyench Sum [ Uench-somon ], coll. 18.06.2007 . Kazakhstan: ZMMU R-8449 (2 juv.) from East Kazakhstan Province, Buran settlement, Chernyi Irtysh, coll. 23.08.1956 ; ZMMU R-10875 (1 adults), East Kazakhstan Province, Kurumcha district, env. of Kamyshinka , left bank of Irtysh River, coll. 05.1956 ; ZMMU R-11989 (2 adults, 1 subad.), East Kazakhstan Province, Aigyrkum sands, 5-7 km SW from Buran, coll. 27.05.2003 ; ZMMU R-14080 (1 adults), East Kazakhstan Province, NE from Ulken-Karatal, coll. 0 8.2013. Almaty Institute of Zoology, Kazakhstan aeae 3420 and 3421 (2 adults), East Kazakhstan Province, Zaisan valley , 10 km S from Ulken-Karatal, coll. 12.07.1985 ; ZIK 3753 View Materials (aeae 22692–22706) (15 adults), East Kazakhstan Province, Zaisan district , 5-6 km N from Karatal, coll. 14- 20.04.1990 .
Diagnosis. A member of the genus Eremias on the basis of the following morphological attributes: ventrals arranged in oblique series, converging at the midline of belly; absence of occipital; subnasal in contact with 1–3 supralabials; large transparent scales on the lower eyelid absent; canthus rostralis not developed (following Szczerbak 1974). A species belonging to the subgenus Pareremias on the basis of the following features: subocular not reaching mouth edge; one frontonasal; two supraoculars; the row of small granular scales between supraoculars and frontal with frontoparietals absent; distance between the femoral pore rows being wide; femoral pore rows not reaching knee-joint; coloration pattern with light colored ocelli with black edging (see Szczerbak 1974). A medium-sized lacertid lizard, maximum snout-vent length (SVL) = 64.5 mm, tail being ca. 1.5 times longer than body length (SVL), hindlimbs relatively long (Pp/SVL ratio 0.46); subocular scale not reaching mouth edge, in touch with 6–8 supralabials; males with bright coloration consisting of 2–3 dorsolateral rows of light-colored ocelli with thick black edging; the ventral row of ocelli in life is greenish to bluish; dorsal pattern consisting of black irregular blotches along the the middorsal line.
Etymology. The specific name “ dzungarica ” is a Latin toponymic adjective in the nominative singular (feminine gender), referring to the distribution of the new species covering the area of Dzungaria, now in easternmost Kazakhstan, northern part of Chinese Xinjiang and the westernmost part of Mongolia (Dzungarian Gobi).
R-12846-2 R-12846-5 R-12846-6 R-12846-7 R-12846-8 R-12846-9 R-12846-10 R-12846-11 R-12862-1 R-12862-2 R-12862-3 Characters m m f juv. juv. juv. juv. juv. m f m SVL 62.5 53.8 64.5 37.0 38.0 34.0 32.0 26.5 60.3 61.4 52.0 Reccomended vernacular name. We recommend the following common name in English: Dzungarian racerunner. Recommended common name in Mongolian: Züüngaryn gürvel; in Russian: Dzhungarskaya yaschurka.
Description of holotype. An adult male preserved in 75% ethyl alcohol in a good state of preservation ( Fig. 3 View FIGURE 3 ); body robust and moderately depressed; a species of the subgenus Pareremias (Szczerbak 1974) ; tail long, ca. 1.5 times longer than body (SVL/TL ratio 0.77). Head relatively long (HL/SVL ratio 0.28) ( Fig. 4 View FIGURE 4 ), massive and wide (HW/HL ratio 0.57), head height slightly less than head width (HH/HW ratio 0.81). Limbs strong, hindlimbs longer than forelimbs (Pa/Pp ratio 0.67), hindlimbs comprise almost half of the body length (Pp/ SVL ratio 0.31). Nostrils not swollen nor raised, three pairs of nasals; subnasal not in contact with rostral shield and is located above first to third supralabials. Snout bluntly rounded, rostral broadly covers the anteriormost part of the dorsal surface of head, but not in contact with frontonasal. Occipital shield absent. Two large nasals and one small nasal in contact with rostral anteriorly, with first, second and third supralabials laterally, and with frontonasal and first loreal posteriorly. Frontonasal single, broader than long, its height less than its length, and laterally frontonasal in contact with first loreal and posteriorly with prefrontals. Two prefrontals contacting each other forming a longitudinal suture; each almost same size as frontonasal, notably longer than broad and laterally in contact with first and second loreals and posteriorly with frontal and a presubocular scale. The single frontal with distinct concavity, slightly bluging, its maximal width in the anterior part ca. twice greater than width in the posterior part. Frontal broadened, much longer than prefrontal and frontonasal together, frontal length ca. two times greater than the distance from its anterior tip to the tip of snout. Frontal anteriorly in contact with prefrontals, posteriorly and laterally in contact with small presupraocular scale and supraoculars, but not separated from the latter by scales, nor by granules, and posteriorly with frontoparietals. Two frontoparietals almost as large as a single supraocular, laterally being in contact with the second supraocular and postsupraocular, and posteriorly with interparietal and parietals. Interparietal small and relatively diamond-shaped with well-pronounced bulging transparent pineal ocellus, surrounded by frontoparietals and parietals; two very large and plate-like parietals, slightly longer than broad, being in contact behind interparietal forming a suture which is ca. two times shorter than interparietal length. Two loreals, first one small, trapezoid-shaped, surrounded by third and fourth supralabials, subnasal, postnasal, frontonasal, prefronal, and the second loreal which is distinctly large and has pentagonal shape. Two supraoculars of same size, anteriorly than the first supraocular a small presupraocular shield of triangular shape, followed by a series of 13–14 granular scales running posteriorly and separating supraoculars from 5–5 supraciliaries. Posteriorly than the second supraocular a postocular shield is present, it’s broader than long, larger than presupraocular and in contact with second supraocular, frontoparietal and parietal medially and with postocular laterally, anteriorly surrounded by several granules; temporal region mostly covered by granular scales becoming larger towards the tympanal region and larger supratemporal shields (9 on each sides), two of them enlarged. Tympanic scale edging the dorsal edge of ear cavity, distinct and elongated obliquely, of semicrescent shape, almost the same size on both sides; tympanum vertically elongated, same size as the orbit. 8–8 supralabials, 6 of which on each side anteriorly to subocular; 6–8 supralabials in contact with subocular which does not border the mouth; the sixth supralabial enlarged of trapezoid shape. Subocular comparatively small, longer than broad with a distinct ridge being extensively in contact with the lower edge of orbit; anteriorly small presubocular shield; posteriorly large postsubocular shield; lower eyelids with a semitranslucent membrane made up of small scales. Gular scales comparatively large, 21 gulars from symphysis of chin shields to median gular, becoming slightly enlarged posteriorly; gular fold well-developed; collar well pronounced, not serratted, made up of 10 enlarged scales, collar scales of uneven size: the medial one the largest, lateral collar scales same size as the adjacent gulars. Five pairs of submaxillary shields, first two pairs in contact, the third to fifth pairs widely separated; the first pair of submaxillary shileds somewhat larger than the fifth and in contact with mental anteriorly, with first and second infralabials laterally; each scale of the second submaxillary pair consists of two fused scales, partially divided by a transverse suture for one third of its width; the fifth submaxillary pair each in contact with the fourth pair anteriorly, being surrounded by 8 granular scales laterally and posteriorly and not in contact with infralabials; 7–7 infralabials. Up to 16 longitudinal and 31 transverse rows of almost squarish pectoral and ventral plates from collar to hindlimbs ( Fig. 3 View FIGURE 3 , B); median ventral wider than long; dorsal scales smooth, granular, of almost same size along the dorsum, juxtaposed; 48 scales across the middle of dorsum, and about 112 scales in a single row from occiput to a point just above the vent; proximal caudals notably larger than posterior dorsals; caudals on dorsal surface of tail complteley smooth, becoming large and elongate distally, arranged in distinct whorls, 24 scales in the 9–10th whorl behind the vent. Upper forearm covered dorsally by enlarged and almost diamond-shaped scales; lower forearm covered with granules; upper hindlimbs covered dorsally by granules, externally by large shields; tibia covered dorsally by granules, ventrally by large smooth plates, three plates in a transverse row; no fringes on the toes, claws light-colored. 22 uni- and bi-carinate lamellae under fourth toe arranged in a single row; 13/12 femoral pores in each side ( Fig. 5 View FIGURE 5 ), distal pores are notably smaller than median ones, almost reaching the knee joint (separated from the joint by a single scale on each side); the two femoral pore series separated anteriorly by a wide space consisting of 8–9 scales; the distance between the femoral pore series is ca. 2 times shorter than length of a single pore series. precloacal region encompassing polygonal shields, the three median ones being the largest; but almost not different from neighbouring scales in size; 6 plates in longitudinal row from the space between femoral pores to anterior edge of the vent.
Measurements and counts of holotype: SVL — 65.0 mm; TL — 85.0 mm; Ga — 32.0 mm; HL — 18.0 mm; HW — 10.3 mm; HH — 8.3 mm; NL — 20.9 mm; Pa — 20.0 mm; Pp — 30.0 mm; HL/SVL — 0.28; HW/HL — 0.57; HH/ HW — 0.81; Pa/SVL — 0.31; Pp /SVL — 0.46. Sq. — 48; G. — 21; Coll. — 11; P.fm. — 13/12; Ventr. — 31; Ventr.Long. — 16; Sq.c.cd. — 24; Lam.subdig. — 22; Lab.min. — 6/6; Infralab. — 7/7; Dist.P.fm. — 2.0; Sq.dors. — 112; Submax. — 5/5, Submax.Cont. — 2 (but the second and third pairs are almost fused).
Coloration and pattern. Scales on head dorsal surface dark grey-brown with unclear pattern of dark-brownish blotches ( Fig. 6 View FIGURE 6 ); smaller dark spots scattered on head sides and in lateral parts of gular area. Ventral surface light beige; in the posterior part of belly — with rare bright yellow and black spots with on the most lateral ventrals. Dark blotches on body flanks forming two regular longitudinal rows. Dorsum uniformly greyish-brown; three longitudinal rows of dark spots in the medial part of dorsum converge in the scapular area forming two longitudinal stripes on the dorsal surface of neck. Two dorsolateral rows of large ocelli lasting from tympanum posteriorly; ocelli in the both rows with think black edging; dorsal row of ocelli whitish to bluish; ventral row of ocelli bluishgreen to grass-green in life. Temporal scales with contrasting pattern (dark brown blotches and light bluish background); posterior edges of upper hindlimbs and forelimbs with a bright greenish spot with black edging. Supralabials and supraoculars with vague dark pattern. The dorsal surface of tail with black spots; proximal upper surface of tail brown, distal part light brownish-grey.
Color in preservative. After 7 years in preservative, the general pattern of the holotype corresponds well to the pattern reported in life ( Fig. 3 View FIGURE 3 ). However warm tints in coloration fade, yellowish spots on ventrals completely disappeared, the ventral row of ocelli appear bluish instead of greenish in life; warmer background coloration of the holotype turned lighter and get bluish tint in preservative.
Variation. For the variation of paratypes see Table 3. The paratypes largely correspond with the description of the holotype (see Fig. 7 View FIGURE 7 ). The juvenile paratypes (ZMMU R-12486-7–11) are much smaller than the adult paratypes (SVL = 26.5–38.0 mm, n = 5; SVL/TL = 0.62–0.70) and notably differ from the other types in coloration pattern (see Fig. 8 View FIGURE 8 ): five interrupted dark nuchal stripes quickly disappear at neck in speckled or reticulated contrasting pattern, in some specimens light interrupted paravertebral stripes discernable, dorsum coffee-brown to dark beige in ground color; head dorsally lightly colored, with two well defined dark brown bands transversely above eyes; dorsolateral rows of ocelli yellowish-green, yellowish coloration also discernable at posterior surfaces of hindlimbs and lateral surfaces of tail; ventral surfaces of tail bright yellow. Despite previously reported morphological differences between highland (Saur and Saikan Mts.) and lowland (Aigyrkum Sands) populations of E. dzungarica sp. nov. from Eastern Kazakhstan (significant differences found in body size and head proportions, as well as in the relative length of hindlimbs, see Orlova 1995), the population from Mt. Ashutas (Eastern Kazakhstan) is not different from lowland Kazakh (Aigyrkum) and Mongolian populations in values of Sq. and also shares similar coloration pattern.
Sexual dimorphism. SVL does not significantly differ among sexes, but females have larger gleno-acetabular distance. Compared to males, females have slender head and body and duller coloration, bright dorsolateral ocelli well-pronounced in males ( Fig. 9 View FIGURE 9 , A) are absent in females ( Fig. 9 View FIGURE 9 , B). The relative head length in males is greater than in females (HL/SVL in males 0.23–0.25 vs. 0.20–0.23 in females).
Geographic variation. Racerunners from Mongolia collected at elevations above 2000 m a.s.l. are characterized by a bright contrasting coloration in males and quite narrow light vertebral stripe without dark pattern. Specimens of E. dzungarica sp. nov. collected in Eastern Kazakhstan (Aigyrkum sands, elevations under 1000 m a.s.l.) are characterized by the larger size of males (SVL up to 71.2 mm) and a quite broad light vertebral stripe; however they are similar to the Mongolian population in scalation characters and general coloration pattern ( Orlova 1995).
Karyotype. The Uyench population of the new species has 2n = 38 (36M + 2m), NF = 38; all chromosomes are acrocentric ( Orlova & Terbish 1997).
Distribution. In Mongolia the “rock form” of multiocellated racerunner which we describe here as E. dzungarica is common in the foothills of Mongolian Altai (Mongolian Dzungaria), in close vicinity of Uyench Sum, Khovd Aimaq ( Orlova & Terbish 1997: Fig. 54) (see Fig. 1 View FIGURE 1 , populations P.17—18). The new species was also reported from the upstreams of the Bulgan-gol River from the elevations above 2000 m a.s.l. We also recorded the new species in Khovd Aimaq, Bulgan Sum, Bayan-Mod, vicinity of Ikher-Toli ( Fig. 1 View FIGURE 1 , population P.16). Outside Mongolia, E. dzungarica sp. nov., previously identified as E. multiocellata , was recorded from Aigyrkum sands in East Kazakhstan Province ( Ananjeva 1972). Later, the new species was recorded in from locality near to Buran settlement, East Kazakhstan Province ( Prokopov 1978). Specimens of the new species deposited in the Zoological institute of Kazakh Academy of Sciences (Almaty) were collected from 20 km NW from Maikapchagai and 10 km SW from Ulken-Karatal ( Brushko & Kubykin 1988; Brushko 1995). Tuniyev & Ananjeva (2010) report this species in Kyzylkum (Kulujun) sands on the left bank of Bukhtarma water reservoir and from Mt. Ashutas, where this species inhabits stony semidesert. The new species is also expected to inhabit the northern part of the Xinjiang Uyghur Autonomous Region north from Tian Shan Mountains (Chinese Dzungaria); the species is assumingly reported for the Chinese herpetofauna as E. multiocellata ( Zhao & Adler 1993) ; however, the precise taxonomic assignation of the Chinese populations would be possible only after a detailed molecular and morphological study.
Ecological notes. In Mongolian Dzungaria, E. dzungarica sp. nov. inhabits elevations up to 2400–2600 m a.s.l., preferring rocky semi-desert or mountain desert habitats with occasional vegetation: Haloxylon (Amaranthaceae) , Caragana (Fabaceae) , Nitraria (Nitrariaceae) , Reaumuria (Tamaricaceae) and Poaceae ( Fig. 10 View FIGURE 10 , A; see Orlova & Terbish 1997, described as the “rock form” of E. multiocellata ). In the vicnity of Uyench Sum, Khovd Aimaq, the new species is found in sympatry with the “sand form” of E. multiocellata sensu stricto (populations P.17 and P.56 respectively, see Appendix II); however the two species appear to be ecologically isolated with E. multiocellata preferring lowland desert habitats with loose sands and dunes at the foothills ( Fig. 10 View FIGURE 10 , B; see Orlova & Terbish 1997, described as the “sand form” of E. multiocellata ), while E. dzungarica sp. nov. inhabits hilly areas, rocky slopes and outcrops and ravines. In Eastern Kazakhstan, however, in Aigyrkum Sands E. dzungarica sp. nov. is found in lower elevations (usually at 400–600 m a.s.l.) and exclusively in sandy areas and on small dunes; whereas at Ashutas Mt. (1000 m a.s.l.) the new species was recorded on clay soils with gravel and occasional rocky outcrops.
Racerunners appear after the hibernation when temperature rises above 18° C (usually, end of April) and are usually active until the end of September — early October ( Orlova & Terbish 1997). Breeding takes place from end of April to early May; the population in Mongolian Dzungaria is reported to have ovoviviparity; from 1 to 4 newborns appear in middle-end of July ( Orlova & Terbish 1997). Before bearing, pregnant females were observed to move to lower sandy areas with denser Artemisia (Asteraceae) vegetation, which probably provide a better habitat for newborns, where they were observed in the same biotopes with the “sand form” of E. multiocellata sensu stricto
In Mongolian Dzungaria, the new species coexists sympatrically with E. arguta and E. multiocellata sensu stricto. No morphological or genetic evidence of hybridization between E. dzungarica sp. nov. and E. multiocellata sensu stricto in the sympatric population P.17 = 56 (see Appendix II) were recorded (9 and 5 specimens examined, respectively).
Comparisons. E. dzungarica sp. nov. is both most morphologically and molecularly similar to the members of E. multiocellata—E. przewalskii species complex from Central and Middle Asia (subgenus Pareremias ); comparison with Eremias species inhabiting this region, as well as with other members of the subgenus Pareremias appear to be the most pertinent.
Differences from sympatric and closely distributed Eremias species. From sympatric E. arguta (subgenus Eremias sensu stricto) the new species is differentiated by smaller size (SVL up to 65.5 vs. maximum SVL 88 mm in E. arguta ), slender body and longer tail (SVL/TL ratio 0.61–0.75 in the new species vs. 0.86–1.26 in E. arguta ), and dorsal pattern (brightly colored ocelli are absent in E. arguta ; in Mongolia this species shows dorsal pattern of large dark transverse spots and blotches on light greyish background color of dorsum).
From E. vermiculata (subgenus Rhabderemias ), also found in south-western Mongolia (but no cases of sympatry with E. dzungarica reported), E. dzungarica sp. nov. can be distinguished by more robust body, shorter tail, presence of bright ocelli on dorsum and body flanks (vs. absent in E. vermiculata ), absence of thin dark vermiculations on dorsum (vs. present in E. vermiculata ) and absence of a complete row of granules, separating supraorbitals from frontal and frontoparietals (vs. present in E. vermiculata ).
From E. argus (subgenus Pareremias ), mostly recorded from central and eastern Mongolia, no cases of sympatry with the new species were reported) the new species can be diagnosed by a more robust body, relatively longer tail (SVL/TL ratio 0.61–0.75 vs. 0.77–1.03 in E. argus ) and longer hindlimbs (Pp/SVL ratio 0.43–0.56 vs 0.38–0.54), dorsal coloration (rows of brightly colored ocelli absent in E. argus ), presence of a signle frontonasal (vs. paired frontonasal in E. argus ).
Differences from other members of the E. multiocellata — E. przewalskii species complex. From sympatric “sand form” of E. multiocellata sensu stricto ( Fig. 11 View FIGURE 11 , A) from localities in vicnity of Uyench Sum (Khovd Aimaq) the new species can be distinguished by larger size (maximal SVL 65.5 mm vs. 57 mm in E. multiocellata sensu stricto), relatively longer tail (SVL/TL ratio 0.61–0.75 in the new species vs. 0.73–0.98 in E. multiocellata sensu stricto) and relatively longer hindlimbs (Pp/SVL ratio 0.43–0.56 in the new species vs. 0.39–0.55 in E. multiocellata sensu stricto). The two species can be also reliably differentiated by dorsal coloration: in E. multiocellata sensu stricto dorsal light sandy background color is covered by sparse small black spots forming irregular longitudinal lines ( Fig. 11 View FIGURE 11 , A–C); males have small light ocelli and a single dorsolateral row of bluish ocelli with thin dark edging (vs. 2–3 rows of large bluish or greenish ocelli with think black edging in the new species). Though subjected to significant geographic variation, most scalation characters in the new species show lower values than in E. multiocellata sensu stricto (Sq. = 43–52, average 47.75 (n = 20) in E. dzungarica sp. nov. vs. Sq. = 50–60, average 54.80 (n = 30) in E. multiocellata sensu stricto; for details see Orlova & Terbish 1997: Table 42).
From E. cf. reticulata (in Mongolia distributed in Gobi-Altai Aimaq between the ranges of E. multiocellata in Dzungar Gobi and E. przewalskii in Trans-Altai Gobi ; see Fig. 11 View FIGURE 11 , J) E. dzungarica sp. nov. can be differentiated by a more robust body with similar body size, shorter tail, smaller number of scales around 9-10th tail ring (21–28 in the new species vs. 23–31 in E. cf. reticulata ) and by dorsal coloration. Though both species have bright ocelli with dark edging, in E. cf. reticulata they are found in both sexes, whereas in E. dzungarica sp. nov. only in males; the dorsal pattern of E. cf. reticulata ( Fig. 11 View FIGURE 11 , J) is resembling that of E. przewalskii ( Fig. 11 View FIGURE 11 , L, M); pterygoid teeth are less developed in the new species, than in E. cf. reticulata (see Orlova & Dunaev 1992).
The allopatric mountain population of Tsagan-Bogd Uul, described as E. m. tsaganbogdensis (Munkhbayar & Borkin 2010) , can be differentiated from the new species by larger body size (maximal SVL 68 mm in E. m. tsaganbogdensis vs. 64.5 in E. dzungarica sp. nov.), comparatively longer hindlimbs Pp/SVL ratio 0.57 ± 0.01 (0.54–0.62; n = 4, males) and 0.56 ± 0.01 (0.53–0.59; n = 6, females) in E. m. tsaganbogdensis vs. 0.52 ± 0.01 (0.48–0.56; n = 10, males) and 0.48 ± 0.01 (0.43–0.50; n = 6, females) in the new species. E. dzungarica sp. nov. has lower number of Sq. = 47.7 ± 0.39 (43–52) vs. Sq. = 54.29 ± 0.63 (47–59) in E. m. tsaganbogdensis , and P.fm. = 11.5 ± 0.23 (9–15) vs. P.fm. = 12.45 ± 0.24 (11–15) in E. m. tsaganbogdensis . Diagnostic characters of scalation include: the fifth submaxillary shield is not divided in the new species (vs. paired in most specimens of E. m. tsaganbogdensis ); dorsal tail scales always smooth in E. dzungarica sp. nov. (vs. tail scales notably keeled in 85% of examined specimens of E. m. tsaganbogdensis , see Munkhbayar & Borkin 2010); and lower number of ventrals 30.7 ± 0.26 (28–32; n = 28) in E. dzungarica sp. nov. (vs. relatively greater number of ventrals 33 ± 0.24; 25–38; n = 10 in E. m. tsaganbogdensis ).
From E. przewalskii ( Fig. 11 View FIGURE 11 , L, M) the new species can be diagnosed by a smaller size (maximal SVL = 65 mm vs. maximal SVL = 84.7 mm for Mongolian and 97 mm for Chinese populations of E. przewalskii ), smaller number of Sq. (43–52 in the new species vs. 47–71 in E. przewalskii ) and smaller number of G. (18–28 in E. dzungarica sp. nov. vs. 24–41 in E. przewalskii ); and by dorsal coloration ( E. przewalskii has reticulated ( Fig. 11 View FIGURE 11 , L) or zebra-like ( Fig. 11 View FIGURE 11 , M) contrasting dorsal pattern and lacks bright ocelli on dorsum). E. przewalskii has welldeveloped pterygoid teeth (1 to 6 teeth on each pterygoid), whereas in E. dzungarica sp. nov. pterygoid teeth are usually absent or may be underdeveloped (see Orlova & Dunaev 1992).
From E. yarkandensis ( Fig. 11 View FIGURE 11 , G, I) the new species can be distinguished by a more robust habitus (vs. comparatively slender head and body in E. yarkandensis ), by subocular being not in contact with mouth edge (vs. subocular shield touching mouth edge in E. yarkandensis ); by having dorsal tail scales always smooth in E. dzungarica sp. nov. (vs. tail scales notably keeled in types of E. yarkandensis ); and by coloration pattern consisting of 3 rows of bright ocelli with thick black edging (vs. occasional small light ocelli in 1-2 dorsolateral rows with thin dark edging in E. yarkandensis ).
From phylogenetically closely related E. buechneri ( Fig. 11 View FIGURE 11 , H) from Kashgaria (southern Xinjiang, China) the new species can be distinguished by much more robust body habitus and massive head (vs. slender body and narrow head in E. buechneri ); by relatively smaller number of Sq. (43–52 in the new species vs. 47–60 in E. buechneri ); by much smaller number of G. (18–28 in E. dzungarica sp. nov. vs. 23–37; average 29.49 in E. buechneri ); and lower number of ventrals 30.7 ± 0.26 (28–32; n = 28) in E. dzungarica sp. nov. (vs. relatively greater number of ventrals 32.2 ± 0.18; 30–36; n = 55 in E. buechneri ); by length of the suture between frontal and frontoparietals being less than the double length of the posterior edge of mental shield (vs. the suture between frontal and frontoparietals being twice the length of the posterior edge of metal shield in E. buechneri ); and by dorsal pattern with contrasting bright ocelli with black edging and dark spots on dorsum in E. dzungarica sp. nov. (vs. uniformly greyish dorsum without clear pattern or ocelli in E. buechneri ).
Genetic differences. The new species is clearly different from all other congeners included in the present phylogenetic analysis with average uncorrected genetic p -distances varying from 3.20% (between E. dzungarica sp. nov. and the E. cf. buechneri from S Xinjiang, China) to 11.10% (between the new species and E. cf. multiocellata from Gansu Province, China) (see Table 2).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eremias dzungarica
| Orlova, Valentina F., Poyarkov, Nikolay A., Chirikova, Marina A., Nazarov, Roman A., Munkhbaatar, Munkhbayar, Munkhbayar, Khorlooghiyn & Terbish, Khayankhyarvagijn 2017 |
Eremias multiocellata
| Gunther Gunther |
Eremias multiocellata yarkandensis
| Blanford 1875 |