
Heliconilla, Dankittipakul & Jocqué & Singtripop, 2012
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3369.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/933EDA4C-B839-FEBD-CBC2-FF22FE113936 |
|
treatment provided by |
Felipe |
|
scientific name |
Heliconilla |
| status |
gen. nov. |
Heliconilla View in CoL View at ENA gen. nov.
Type species. Heliconilla thaleri ( Dankittipakul & Schwendinger, 2009) View in CoL comb. nov. Heliconilla irrorata View in CoL comb. nov. was described by Thorell (1887) and is the oldest description of a species belonging in the genus. As it was based on an immature specimen, we decided not to select it as the type species.
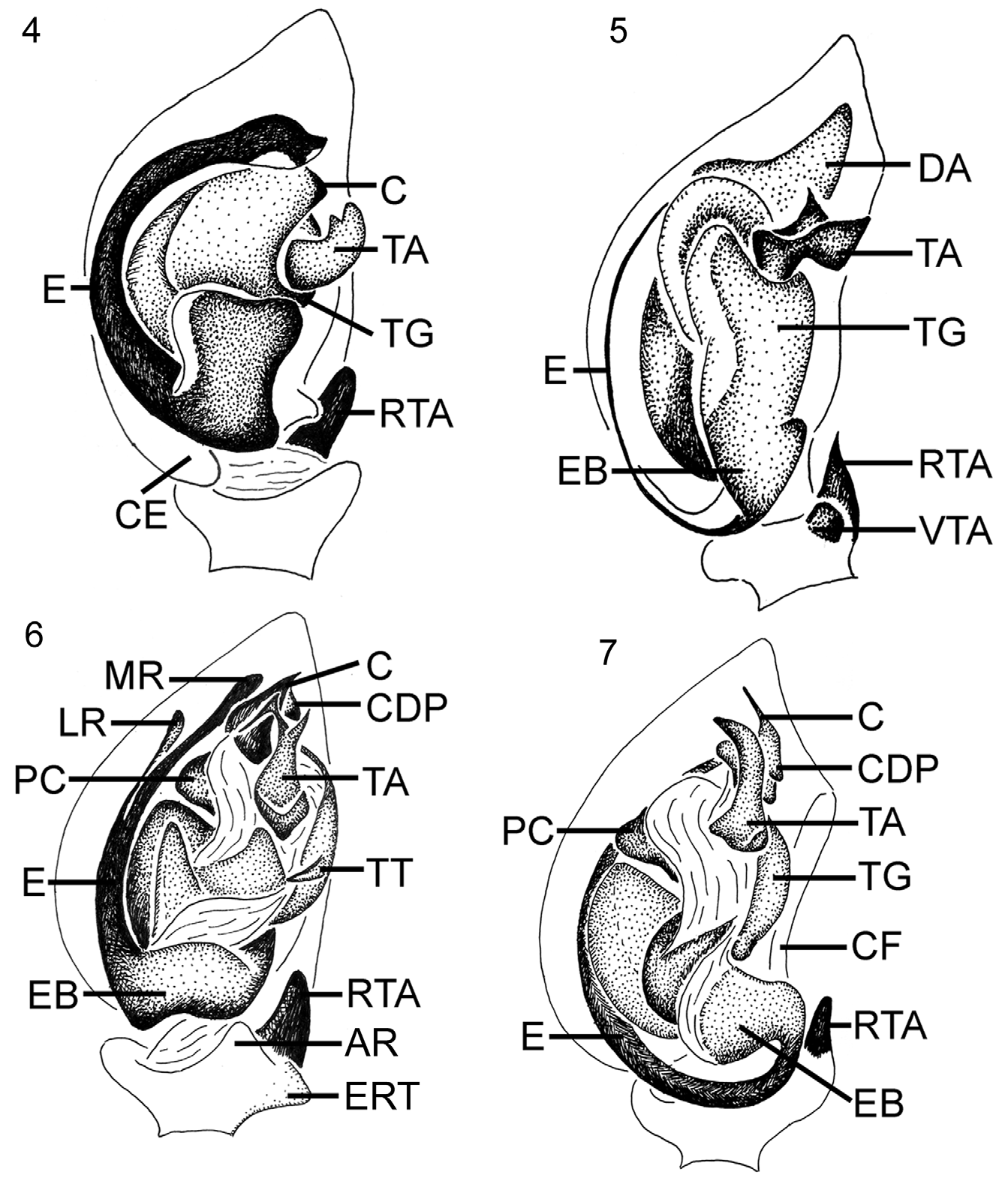
Diagnosis. Members of Heliconilla gen. nov. can be recognized by a combination of eleven synapomorphies (Ch. 4–5, 18–21, 24, 37, 58, 62, 80) and five homoplasious characters (Ch. 50, 54–55, 73, 89). The convex carapace is strongly granulate in both sexes, covered with brachiate setae on elevated sockets (Ch. 5) (Figs 1285–1288). The posterior ventral spines are represented by a group of erect, modified setae situated on a chitinized plate; these spines are short and bluntly pointed in males (Fig. 1292), thin and hair-like in females (Fig. 1291); their apical surface is serrated, provided with two parallel rows of minute denticles (Ch. 24). The cymbium on the male palp is thickly covered by elongated chemoreceptive and brachiate hairs (Ch. 58) (Figs 1293–1296). The terminally flattened apex of the conductor is extended anteriorly; the dorsal process of the conductor is blunt and lobular ( Figs 7 View FIGURES 4–7. 4–7 , 1297–1298). The sickle-shaped TA is simplified and partially membranous, always provided with a basal tooth and a lightly sclerotized apico-prolateral flange situated anterior to a membranous area (Ch. 80) ( Fig. 38 View FIGURES 25–38. 25–33, 35–37 ). The embolus is generally blade-like, frequently with an elevated median ridge running along its entire length (Ch. 73), and usually furnished with an accessory process (Figs 1317–1322). The trapezoidal epigynal plate lacks a prominent anterior median incision; its posterior margin is generally protruding over an epigastric furrow (Fig. 1281). The internal genitalia are characterized by short, triangular insemination ducts; the spermathecae consist of an elongated proximal part provided with a posterior lateral swelling and a strongly convoluted distal part usually forming globular bulges (Figs 1309–1315). The dorsal pattern on the opisthosoma is uniform, represented by a median longitudinal band of white pubescence in the live spider ( Fig. 95 View FIGURES 92–95. 92 ); this pattern is replaced by a pair of pale markings, followed by an irregular arrangement of minute pale spots in preserved specimens (Ch. 18–20) (Figs 1271, 1273); the venter is mottled with numerous pale spots on dark sepia background (Ch. 21) (Figs 1272, 1274). The male dorsal scutum is lanceolate.
Etymology. The genus name is an arbitrary combination of letters. The gender is feminine.
Affinities. Heliconilla gen. nov. is an easily recognized monophyletic genus united by several synapomorphic characters which seem to be apomorphic within the Zodariinae . Ancestral Zodariidae have simple, aculeate setae covering the ventral side of the opisthosoma. In some genera of the subfamily Zodariinae (e.g. Workmania gen. nov., Mallinella , Euryeidon , Heradion ), these setae are significantly enlarged and sit on a clearly defined chitinized base ( Figs 8–11 View FIGURES 8–11. 8, 10 ). Heliconilla gen. nov. is also characterized by the extraordinary modification of these setae which have become the posterior ventral spines (PVS). The presence of PVS is not unique to Heliconilla gen. nov., being found in both sexes in a few other genera mentioned above, but the surface of these spines is generally smooth and simple. The microstructure of the PVS in Heliconilla gen. nov. is modified, their surface is serrated, provided with two rows of denticles (Figs 1289–1292). This character is unique and is one of the synapomorphies.
Apart from the modification of the PVS, some characters are not present in any other taxa. All species appear to have a strongly granulated carapace which is covered with micro-protuberances, each consisting of 15–25 mosaic plates (Figs 1285-1288). Generally, the protuberance on the carapace of other zodariids is not modified and represented by a single, elevated mosaic ( Figs 334–335 View FIGURES 332–335. 332–335 ).
The presence of brachiate setae on the carapace is of interest. In other zodariids, the setae on the carapace are simple, aculeate and sparsely distributed ( Figs 175 View FIGURES 174–177. 174–177 , 939 View FIGURES 936–941. 936–941 ), whereas in Heliconilla gen. nov. the brachiate setae are numerous (Figs 1287–1288). Together they form a distinctive patch that is noticeable in the live spider ( Fig. 95 View FIGURES 92–95. 92 ). This broad median patch runs longitudinally between the ocular region and the posterior margin of the carapace ( Fig. 95 View FIGURES 92–95. 92 ). The peculiar pattern continues on the opisthosoma where it is visible as a longitudinal band of white pubescence on the dark opisthosomal background. Unfortunately, this pattern is unclear in the preserved specimens, where only the first and second pairs of pale markings remain (Figs 1297-1308). In Mallinella , the venter of the opisthosoma is marked with a broad longitudinal band, flanked by two oblique lateral bands on pale background ( Fig. 721 View FIGURES 719–725. 719-722, 724-725 ), whereas in all members of Heliconilla gen. nov. the background is dark and asymmetrically mottled by numerous pale spots (Figs 1272, 1274).
The male palpal morphology of Heliconilla gen. nov. is comparatively simple and uniform in all species, showing no extraordinary diversity, except little variation is present in association with the embolus, which is considered species specific. The presence of the modified embolus is possibly associated with the loss of the necessity to produce a complex TA (in contrast to Mallinella species , whose TA is of variable shapes and sizes). An apex of the conductor is elongate, extending anteriorly and terminally flattened while in Mallinella species , it is sharply pointed. A dorsal process of the conductor is not modified (triangular or digitiform in Mallinella ), represented by a blunt lobe. The presence of numerous brachiate cymbial setae and distinctly elongated chemoreceptive hairs on the dorsal side of the cymbium has been confirmed in all species, and the microstructure of these hairs is identical in all species examined. This makes the male palp of live spiders appears sticky and it is usually encrusted with soil and litter particles. The modification of these hairs is of interest and common among species of Heliconilla gen. nov., unfortunately, their taxonomic value cannot be established at this stage because the information lacks for most representatives of other genera.
Although the heavily sclerotized epigynal plate is almost similar in all species, represented by a simple, trapezoidal plate, the female internal genitalia show extraordinary diversity at the species level. Near the base of each spermatheca is a prominent semi-circular swelling of unknown function. The basal part of the spermathecae is generally elongate, whereas their distal part is convoluted and modified, forming a globular apical bulge. This conformation appears to be the autapomorphic condition for the genus.
Description. Medium-sized to large zodariid spiders. Prosoma elongate-ovoid (Fig. 1285), widest at level between coxae II and III, pars cephalica strongly convex, in profile highest just in front of fovea, gradually sloping backwards; integument strongly granulated, each hump consisting of 15–25 micro-mosaic plates (Figs 1286–1288). Carapace uniform dark brown, usually with shallow longitudinal furrow running between PME and deep, longitudinal fovea; always covered with brachiate setae situated on distinct sockets (Fig. 1288); longer aculeate setae sparsely distributed, denser in ocular region. Chilum single sclerite, triangular, elevated, covered with few setae. Chelicerae with distomesal group of hairs; fangs short and stout; no denticles on cheliceral margins; lateral condyles moderately elevated. Labium longer than wide, slightly constricted at base, sparsely covered with setae. Gnathocoxae triangular, covered with black anteromesal scopulae. Sternum scutiform (Figs 1272, 1274), medially elevated, anterior margin straight, lateral margin with small, triangular extensions fitting into coxal and intercoxal concavities; sigilla shallow, situated on posterior margin. All eyes circular, pale (Figs 1275–1276); AER and PER strongly procurved; AME enlarged, less than radius apart, at about 1.5 diameters from ALE; PME about 1 to 1.5 diameters apart and about 2 to 2.5 diameters from PLE. MOQ longer than wide; distinctly wider in front than behind. Leg formula 4123. Legs with erect spines on tibiae, metatarsi and tarsi, more numerous spines on posterior legs; ventral surface of metatarsi with thick brush of chisel-shaped distal setae, setae distinctly denser on posterior legs. Opisthosoma elongate-ovoid. Dorsum of opisthosoma with distinct dorsal pattern: anteriorly with a pair of <> markings, followed by a pair of oblique bands and by a series of short irregular bands, together with numerous pale spots (Figs 1297–1308). Dorsal scutum lanceolate, lightly sclerotized, occupying approximately 1/3 length of opisthosoma. Epiandrum present. PVS (Figs 1289–1292) erected, arranged into a group on sclerotized plate, bluntly pointed in males, thin and hair-like in females; surface of PVS serrated, provided with two rows of minute denticles. Spinnerets as usual in the subfamily Zodariinae .
Palp ( Fig. 7 View FIGURES 4–7. 4–7 ). RTA digitiform, broad at base, gradually tapered towards bluntly pointed apex. Second RTA (also known as DTA) minute and triangular, situated above RTA (Fig. 1293). Cymbium with narrow basal fold occupying 2/3 of cymbial length or almost reaching apex. TA sickle-shaped, broad at base, subterminally abruptly bent mesad, always with lightly sclerotized apico-prolateral flange and basal ridge or tooth. Tegular spine absent. Conductor elongated, with flattened apex directed anteriad; dorsal process of conductor short, terminally blunt. Embolus blade-like, bifurcated (usually deviating from base), or with accessory process.
Female. Generally as in males but carapace more elongated, anteriorly slightly wider; dorsal scutum absent.
Genitalia. Epigynal plate subrectangular, anterior margin straight or slightly excavated (but never with anterior median incision), posterior margin shorter, usually protruding over epigastric furrow. Internal genitalia heavily sclerotized, with short, diverging triangular insemination ducts. Spermathecae consisting of elongated proximal part with posterior lateral swelling and strongly convoluted distal part.
Species included. Nine species: Heliconilla aculeata sp. nov., H. cochleata sp. nov., H. crassa sp. nov., H. furcata sp. nov., H. globularis sp. nov., H. irrorata ( Thorell, 1887) comb. nov., H. oblonga ( Zhang & Zhu, 2009) comb. nov., H. mesopetala sp. nov. H. thaleri ( Dankittipakul & Schwendinger, 2009) comb. nov.
Natural history. Species of Heliconilla gen. nov. are generally nocturnal and live in lowland rainforest to mountain forests with constant high humidity, only a single species was found in more open habitats, such as deciduous dipterocarp forest, and in degenerated secondary forest ( H. oblonga comb. nov.).
Distribution. Indo-Burma and Sundaland subregions of the Oriental Region (southern China, Thailand, Burma, Vietnam and Malaysia).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |