
Sibogaster nieseni
|
publication ID |
https://doi.org/10.11646/zootaxa.4175.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:B264C215-000D-42C5-8AC9-B801872CD182 |
|
DOI |
https://doi.org/10.5281/zenodo.6055262 |
|
persistent identifier |
https://treatment.plazi.org/id/953787B9-FF90-FFCC-5CF0-FDD181C7FA88 |
|
treatment provided by |
Plazi |
|
scientific name |
Sibogaster nieseni |
| status |
|
Sibogaster nieseni View in CoL nov. sp.
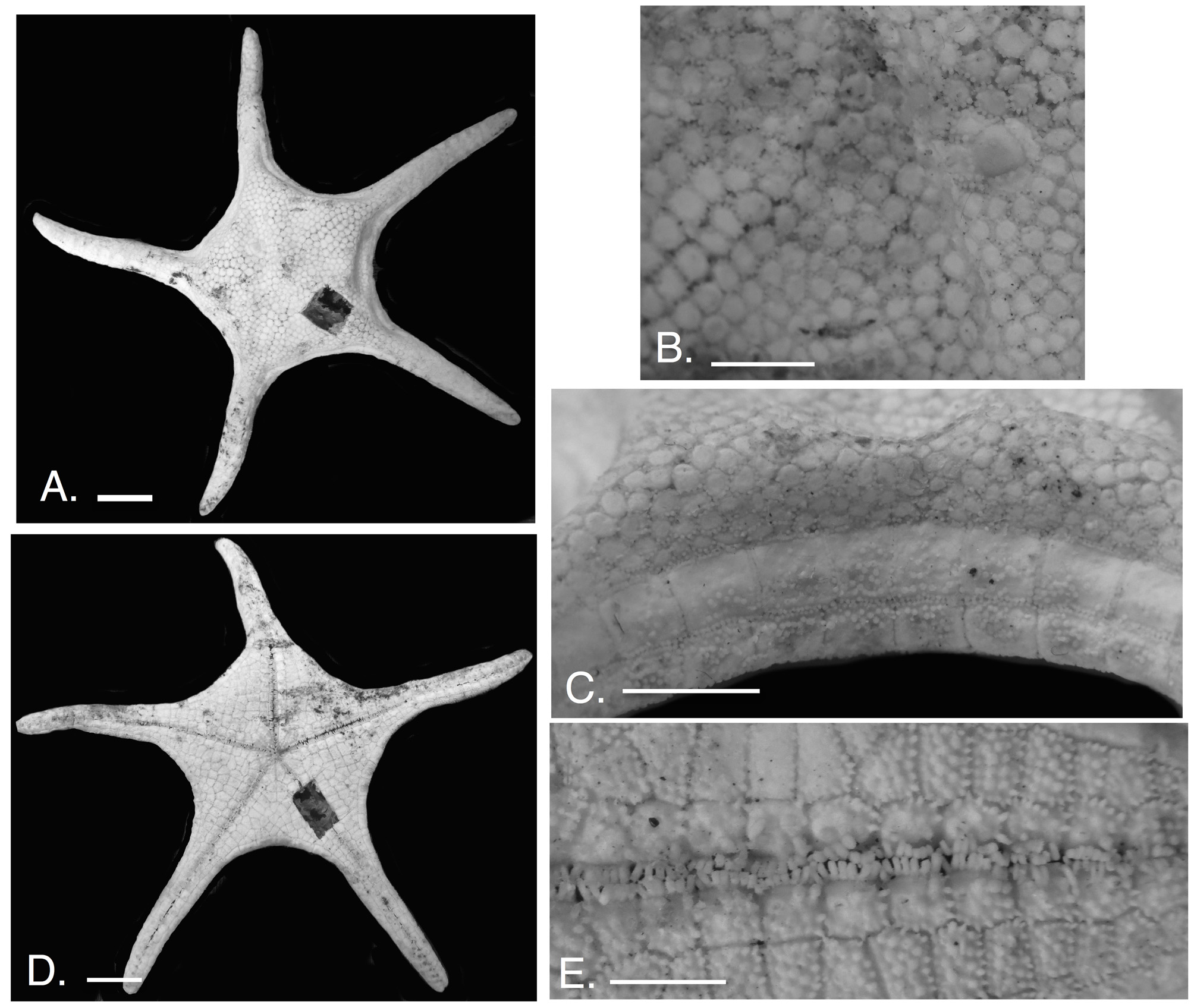
Figure 11 View FIGURE 11 A–E (North Pacific), 12A–E ( Indonesia), 13A–E (Atlantic)
Luke 1982: 15; Maluf 1988: 34, 119 (as cf. Sibogaster sp.)
Etymology. This species is named for Professor Thomas Niesen, Professor Emeritus of Marine Biology, San Francisco State University.
Comments. Specimens of Sibogaster nieseni n. sp. were first brought to the attention of the author via specimens collected by MBARI, but as museum collections were reviewed, it was determined that this specimens of this species had been collected as early as 1966 ( Luke 1982: 15) from collection in the Pacific and the Atlantic.
This species was placed into the genus Sibogaster based on the shared smooth abactinal and marginal plate surfaces as well as shared actinal granulation and similarity in furrow spine number and morphology. As indicated below, specimens from several widely occurring localities in the Pacific and the Atlantic were identified. Morphological differences between individuals was slight, including similar to identical numbers of furrow spines (six to six or eight), actinal granules ranging from round to round with pointed tip, to minor variation in presence/ absence of pedicellariae on marginal plates. Other characters were mostly identical, abactinal and marginal plates were bare with coarse granules between them and body shape was largely similar. Differences were similar to those observed in other widely occurring species, such as Hippasteria phrygiana (Mah et al. 2014) or in species of the deep-sea Porcellanasteridae (e.g. Madsen, 1961a).
The Indonesian specimen (IE-2013-6809) shows perhaps the most significant differences relative to the other specimens in that the abactinal accessory and actinal granules are pointed and spinelet-like. Also dissimilar is the presence of granules on several of the interradial marginal plates which then trail off in number, the plates becoming bare along the arms.
This species is unusual for the Goniasteridae in that it occurs well below 2000 m (lower Bathyal zone). Most goniasterids occur either on the continental shelf or the upper bathyal zone ( Mah & Blake 2012) with some taxa occurring in deeper-waters at their lower range (e.g., Plinthaster , Anthenoides , etc.) and only a few others have this depth as their primary occurrence (e.g., Pillsburiaster ).
In situ Observations. In situ observations of this species from Taney Seamounts B and C as well as the Gorda north Escanaba Trough (aka the NESCA clam site) in the North Pacific revealed that this species displayed an inflated disk. Based on the distended abactinal disk surface of the preserved specimen and the absence of sediment in the gut, the coelom was swollen with seawater. These observations showed S. nieseni on fine mud. Although stomach dissections were inconclusive, some mud was present in the oral/stomach region and along the tube feet suggesting deposit feeding. The cardiac stomach in this species appears well-developed.
The specimen collected from the NESCA clam site (USNM 1407956) was within the influence of a cold seep site with calyptogenid bivalve shells ( Calyptogena extenta ) sitting nearby. Other megafauna observed in close proximity included Tonrometra (Crinoidea) , Synallactidae (Holothuroidea) , Brisingida (Asteroidea), Benthothuria and Paelopatides confundens (Holothuroidea) , Actiniaria, and Munidopsis sp.(Crustacea).
The specimen from Taney seamount B (USNM 1407955 D173-A7) were collected from the thin sediment ponds between large pillow lavas. Other fauna observed included isidid octocorals, various holothurians, crinoids ( Pentametrocrinus paucispinulus and Tonrometra ), several sponges, and Xenophyophoroidea. USNM 1407957 (D178-A21) was collected in proximity to several species of sponges ( Aspectopluma sp., Caulophacus sp.) Tonrometra (Crinoidea) , Calliostoma sp. (Gastropoda), ophiuroids, and Heteropolypus sp. ( Alcyoniidae, Octocorallia ).
Occurrence. North & Central Pacific. Taney Seamounts B and C, “ NESCA ” Clam sites ( North Pacific ), Patton Seamount and Patton Escarpment , Cascadia Plain, west of Cortez Bank , Celebes Sea, Indonesia, 2100–4175 m . Tropical Atlantic , off coast of Mauritania, 2114–3162 m . Combined Depth Occurrence : 2100–4175 m .
Description. Body stellate (R:r=1.88 to 3.0, Atlantic specimens 2.0–27.7), arms elongate but triangular in shape, abactinal disk surface distended. Interradial angles weakly curved to linear ( Figs. 11 View FIGURE 11 A, 12A, 13A).
Abactinal plate surface smooth, devoid of accessories (no surficial granules, etc.) ( Fig. 11 View FIGURE 11 B,C; 12B;13B,C). Plate surface flattened (in NESCA specimen) and level with marginal plate boundary. On some specimens (e.g., CASIZ 118414) the disk is more swollen and rises above the plane of the marginal plates. Most plates flat but some with weakly convex surface; these occur more distally along arm, near terminus and in some specimens adjacent to superomarginal plate surface. Outline shape round/polygonal to oblong, plates larger proximally becoming smaller, more irregular in shape and more abundant closer to contact with superomarginal series. Although a majority of proximal plates are relatively large (~ 0.8 to 1.2 mm diameter) a number of smaller ( 0.5 to 0.9 mm diameter) plates are also present. Carinal series is present but not distinctive. Shallow grooves present along radial regions, absent interradially. Abactinal plates irregularly arranged, more abundant proximally declining to a single row (the carinal series) distally on arm, extending to armtip. Each plate periphery surrounded by discrete ring of granules, 10–35 (usually 20–25), each oval shaped to more quadrate in outline. Peripheral granules proximally large, taking up to 25–30% of the abactinal plate plus granule diameter. Size of these granules becomes smaller distally along arm, becoming less prominent on highly convex distal plates. Peripheral granules are variably distributed, with some closely arranged versus others showing granules more discontinuously arranged.
Papulae large, three to six per plate, occurring at interstices between other adjoining abactinal plates, present only on radial regions over arms, absent interradially. Madreporite, flat to weakly convex, round to pentagonal in outline, flanked by five to fifteen adjacent abactinal plates. Pedicellariae, small (~ 0.25 mm), when present, tonglike with blunt, paddle-like blades and inset within a shallow pit. Pedicellariae are absent in some specimens but not in others, primarily occurring proximally on disk.
Marginal plates wide (distally) to quadrate (interradially), facing laterally, superomarginal and inferomarginal plate series with 1:1 correspondence interradially but becoming offset distally. Marginal plates, flush with body surface, closely articulated ( Fig. 11 View FIGURE 11 B; 12C; 13C, D). Fasciolar grooves absent. Marginal plates surfaces largely bare of accessories. Superomarginal plates, 28–46, (mostly 34–38 at ~R=4.0–5.0 cm, but R=3.4, USNM 1407957 with 30 superomarginals) from armtip to armtip, squarish in outline with length and width nearly equal interradially with distinct angular edges, becoming more rounded distally along arms. Superomarginal surface bare, flattened interradially becoming more strongly convex distally. A minority of superomarginal plates with no other accessories except for one to five round, hemispherical granules on two to five interradial plates. The Indonesian specimen ( Fig. 12 View FIGURE 12 A,B) with more abundant granular cover on the marginal plates interradially becoming bare distally. Similar in outline to superomarginals but having more convex to round facing along contact with actinal surface. Inferomarginal plates, 28–46, surface flattened interradially becoming more round and convex distally near terminal plate. Plate surface bare on upper side adjacent to superomarginal series, but covered by granules, 7 to 26, on lower side of the inferomarginal plates in contact with inferomarginal series (in addition to the lower peripheral granule series). These granules most abundant interradially but absent distally; interradial plates are similar in size and density to those present on the actinal surface and may completely cover up the lower inferomarginal plate surface. In Pacific specimens, the granules are more similar in size, in Atlantic ( Fig. 13 View FIGURE 13 C) specimens, the inferomarginal granules are slightly coarser than those on the actinal surface. Peripheral granules 40–70 (usually about 60) at R=~5.0; granules are similar to those on abactinal plates present, surrounding each plate, these are large proximally becoming smaller distally. Pedicellariae small, one or two per plate, similar to those present on the abactinal surface, variably located on a minority of marginal plates. Terminal plate large, round, bare with nipple-like protuberance present.
Actinal intermediate region large, composed of approximately four to six distinct chevrons ( Figs. 11 View FIGURE 11 D, 13E, F). Actinal plates ranging in shape from quadrate, hexagonal to irregularly polygonal adjacent to the inferomarginal contact. Outermost chevron adjacent to the adambulacral plate series is distinct but actinal intermediate plate series become increasingly more irregular closer to inferomarginal plate contact. Actinal plates extend from disk to approximately 50% of arm distance. Surface flat, each with 8–30 hemispherical to pointed granules per plate forming a dense covering over the complete actinal surface. Peripheral granules range10–25 with six to 15 present on plate surface (these latter granules are larger than those that compose the periphery). Granule arrangement variable with some individuals showing densely crowded, more swollen granules in contrast with other individuals with more widely spaced, relatively smaller granules. Granule distribution overall is more widely spaced proximally becoming more densely packed distally adjacent to the inferomarginal contact. On both Pacific and Atlantic individuals, granules are round to pointed. In some Pacific individuals, such as the Indonesian specimen ( Fig. 12 View FIGURE 12 B, IE-2013–6809, R= 3.5 cm) granules are acutely pointed and are nearly spinelet-like. Pedicellariae observed on only a one or two plates on actinal region and only on a minority of Atlantic and Pacific specimens. Valves are flattened, paddle-shaped, sitting within an inset, present on proximal edge of actinal plates near oral region.
Furrow spines six to ten, mostly seven to eight increasing distally with largest number of furrow spines in largest specimens ( Figs 11 View FIGURE 11 E, 12E, 13F). Furrow spines mostly quadrate in cross-section. Some specimens (MNHN IE-2013-8190) show more flattened furrow spines versus others which display more thickened to quadrate spines in cross-section (e.g., MNHN IE-2013-8194. Most furrow spine tips blunt (e.g, CASIZ 120367) but one with notching on tips (USNM 1407956, NESCA clam site) was observed. A discrete space separates furrow spines from subambulacral accessories (granules, spines, etc.). Subambulacral spines three to five, round to polygonal in crosssection. In those with three, two distalmost spines twice as thick as furrow spines and third proximal spine as thick as but approximately half the length of furrow spines. In those with five, all subambulacrals identical in size and thickness but of paddle-like shape. In some Atlantic individuals (MNHN IE-2013-8190), subambulacral spines more triangular to oval in outline with pointed tips, about 10% thicker than furrow spines. These spines are variably widely spaced or arranged close together. Adambulacral pedicellariae one or two, elongate, tong-like in shape, thick with complementing valves, taller than furrow spines. Pedicellariae, variably present on adambulacral plates sitting adjacent to subambulacral spines, occurring irregularly from the third adambulacral and distally along the furrow series.
Oral plates with 8 to 10 furrow spines, quadrate to triangular in cross-section with one large flattened spine per plate, directed into the mouth (so two total per interradius projecting into the mouth.). Oral plate surface covered by 8 to 15 enlarged prominent granules, quadrate to triangular in cross-section, sometimes closely articulated next to one another in series ( Fig. 11 View FIGURE 11 E, 13F). The enlarged granules are paired off on each oral plate present interradially forming a cavity between the two series. Five to ten similar granules are present on each of the oral plates. Discrete spaces are present between the furrow spines and subsequent granules. Other than these granules, oral plates are bare.
Color in life is orange to deep orange-red.
Material. HOLOTYPE USNM 1407955, Taney Seamount, 36.774013, -125.363497, 3052.5 m, Coll. Dave Clague et al., MBARI-ROV, D173-A7. 1 dry spec. R=4.7, r=1.6.
PARATYPES: North Pacific. USNM 1407956 About USNM “ NESCA clam site”. 41° 0' 44.2 N, 127° 29' 28.3 W GoogleMaps , 3321. 8 m. Coll. Dave Clague et al, MBARI-ROV Doc Ricketts, D83-A6. 1 wet spec. R=6.0 r=3.0 cm. USNM 1407957. Taney Seamount C. 36° 42' 55.6, 125°N, 14' 52.9 W 3283.1 m. Coll. Dave Clague et al, MBARI-ROV Doc Ricketts, D178-A21, 1 wet spec. R=3.4, r=1.5. CASIZ 118414 . Off the Oregon coast. 44°53.5’N, 127°27.5’W GoogleMaps , 2826 m. Coll. R/V Cayuse, Cr. 7310B (BMT 332), 4 Nov. 1973. 2 wet specs. R=5.1, r=2.6, R=4.9, r=2.6; CASIZ 120367 . Patton Seamount , 54.600,-150.468 to 54. 570, -150.485, substrate was 75% sand, 25% cobble , 2080–2732 m. Coll. Jerry Hoff aboard R/V Alvin, 25 July 1999. 1 wet spec. R=5.1, r=2.1; CASIZ 121402. Cascadia Plain. 44°33’N, 128°19.2’W, 2820 m. Coll. R/V. Cayuse. Cr. 7310B (BMT 331), 3 Nov. 1973, 1 wet spec. R=6.4, r=2.9; SIO E 308. WSW of Cortez Bank. 32°05’N, 120°29’W to 32°03’N, 120°30’W, 3777–3792 m. Coll. C. Hubbs, MV 66-III-21. Dec. 11, 1 966, 1 wet spec. R=3.7, r=1.6; SIO E 1551. W. of Patton Escarpment. 32°22’N, 120°42’W, 3640–3740 m, Coll. R / V N. Horizon, K. Smith & S. Luke, Feb. 6, 1981, MET St. 129, 1 wet spec. R=5.6, r=2.5; Tropical Pacific GoogleMaps . MNHN IE- 2013-6809 . Celebes Sea, Indonesia. 5º7’N, 125º 9’E, 4700– 4175 m, Coll. ESTASTE II expedition, Trawl 10, 12 Jan, 1984. 1 dry spec. R=4.9, r=1.6. Tropical Atlantic. MNHN-IE-2013-8188 Tropical North Atlantic , off coast of Mauritania, 18˚14.64’W, 20˚33.66’W GoogleMaps , 3160 m, Coll. M. Sibuet, EUMELI 2, CPH 13, 24 Jan 1991. 1 dry spec. R=3.1, r=1.2; MNHN-IE-2013-8189 Tropical North Atlantic , off coast of Mauritania, 18˚14.64’W, 20˚33.66’W , 3162 m. Coll. M. Sibuet, EUMELI 2, CPH 2, 27 Jan 1991, 1 dry spec. R=6.1, r=2.4. MNHN-IE-2013-8190 Tropical North Atlantic , off coast of Mauritania. 18º14.64’N, 20º 33.66 W GoogleMaps , 3162 m. Coll. M. Sibuet, IFREMER, N/O L’Atlante, CENTOB CPH 9 About CPH . 11 Feb. 1991. 1 dry spec. R=4.7, r=2.1. MNHN-IE-2013- 8191 Tropical North Atlantic , off west coast of Mauritania. 20º17.04”N 18º 2.76’W, 2114 m. Coll. M. Sibuet et al GoogleMaps . IFREMER aboard N/O L’Atlante, EUMELI 2, CPH 3, 5 Feb. 1991. 1 dry spec. R=5.2, r=1.8. MNHN-IE-2013- 8192 NW Atlantic. Barbados. 10º19.97’N, 58º 37.30 W GoogleMaps , 2924 m. Coll. K. Olu, 1 May 1970 aboard S/ M Nautile (N/ O Nadir), DIAPISUB, Pl. 10-16, 1 dry spec. R=3.1, r=1.2. MNHN-IE-2013-8193 Tropical Atlantic , off coast of Mauritania. 18º 14.64 N, 20º 33.66 W GoogleMaps , 3162 m, Coll. M. Sibuet, aboard N/O L’ Atllante, 1 dry spec. R=5.0, r=1.8. MNHN-IE-2013-8194 Tropical Atlantic, off coast of Mauritania. 18º 14.64 N, 20º 33.66 W GoogleMaps , 3162 m. Coll. M. Sibuet, IFREMER, N/O L’Atlante, EUMELI 2, CPH 1. 24 Jan. 1991. 1 dry spec. R=5.2, r=2.1.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |