
Setaphyes elenae, Cepeda & González-Casarrubios & Sánchez & Pardos, 2020
|
publication ID |
https://doi.org/10.5852/ejt.2020.637 |
|
publication LSID |
lsid:zoobank.org:pub:BD1FDE40-F1C4-49C8-BCDB-72AB3EDA9F17 |
|
DOI |
https://doi.org/10.5281/zenodo.3795356 |
|
persistent identifier |
https://treatment.plazi.org/id/6C4FE50F-E39A-451E-B805-60B709B9B9B4 |
|
taxon LSID |
lsid:zoobank.org:act:6C4FE50F-E39A-451E-B805-60B709B9B9B4 |
|
treatment provided by |
Valdenar |
|
scientific name |
Setaphyes elenae |
| status |
sp. nov. |
Setaphyes elenae sp. nov.
urn:lsid:zoobank.org:act:
Figs 2–4 View Fig View Fig View Fig , Tables 1–3 View Table 1 View Table 2 View Table 3
Diagnosis
Setaphyes with shortened, distally rounded middorsal processes on segments 1 and 9, and middorsal elevations on segments 2–9, superficially covered by tufts of elongated, thick hairs whose tips sometimes surpass the posterior margin of segment. Unpaired setae in paradorsal position on segments 1–9. Laterodorsal setae on segments 3, 5, 7 and 9; paralateral setae absent. Lateroventral setae on segments 2–10. Ventromedial setae on segments 3, 5 and 7. Paired, small, dot-shaped intracuticular structures (maybe outlets of glandular cells) present in several positions throughout the trunk, with a specific arrangement that differs from males to females. Males with paired, sexually dimorphic ventromedial tubes on segment 2, and females with paired, sexually dimorphic ventrolateral setae on segment 2. Lateral terminal spines present, relatively short, slender. Segment 11 retractable into segment 10.
Etymology
The species is dedicated to Ms Elena González, sister of the second author.
Material examined
Holotype
ATLANTIC OCEAN • ♀ adult (mounted in Fluoromount G ®); near Syd-Hällsö Island, Skagerrak ( Fig. 1B View Fig ); 58°56.846′ N, 11°4.896′ E; 55–65 m depth; Ulf Jondelius and Fredrik Pleijel leg.; very fine mud; NHMD 655358 .
GoogleMapsParatypes
ATLANTIC OCEAN • 3 adult ♂♂, 2 adult ♀♀ (all mounted in Fluoromount G ®); same collection data as for holotype; NHMD 655359 to 655363 .
Additional non-type material
ATLANTIC OCEAN • 8 specs (four mounted for LM and four mounted for SEM); same collection data as for holotype; Meiofauna Collection UCM.
Description
See Table 1 View Table 1 for measurements and dimensions, Table 2 View Table 2 for summary of cuticular elevation, process, seta, tube, nephridiopore and sensory spot locations, and Table 3 View Table 3 for summary of intracuticular, dot-shaped structure locations.
Head with retractable mouth cone and introvert. The collected specimens were not suitable for head examinations, hence data on number and arrangement of scalids and oral styles are not available.
Neck with four dorsal and two ventral sclerotized placids ( Fig. 2 View Fig A–B, D). Dorsal placids rectangular, with a slightly convex anterior margin; mesial ones broader than lateral ones ( Fig. 2B View Fig ). Ventral placids morphologically similar to dorsal ones but much more elongated, getting thinner towards the lateral sides ( Fig. 2A, D View Fig ).
Trunk with eleven segments ( Figs 2 View Fig A–B, 3A, H, 4A). Segment 1 with one tergal, two episternal and one sub-trapezoidal, midsternal plate; remaining ones with one tergal and two sternal cuticular plates ( Figs 2 View Fig A–D, 3A, H). Tergal cuticular plates slightly bulging middorsally. Sternal plates reach their maximum width at segment 5, but are almost constant in width across the trunk, slightly tapering at the last three trunk segments ( Figs 2 View Fig A–B, 3A, H). Sternal cuticular plates are relatively narrow in the ratio maximum width to total trunk length (MSW-5: TL average ratio = 27.2%), giving the animal a slender appearance ( Figs 2 View Fig A–B, 3A, H, 4A). Middorsal processes on segments 1 and 9, shortened and distally rounded ( Figs 2B View Fig , 3B, M View Fig , 4G, I); middorsal elevations on segments 2–8, pentagonally-shaped, distally rounded, with intracuticular, butterfly-like atria of paradorsal sensory spots ( Figs 2B View Fig , 3D, F, I, K View Fig , 4B, E). Middorsal elevations superficially covered by tufts of elongated, thick cuticular hairs whose tips sometimes surpass the posterior margin of the segments ( Fig. 4B, E View Fig ). Intracuticular, minute, dot-shaped, rounded to oval structures (maybe outlets of glandular cells) throughout the cuticle on segments 1–10 ( Figs 2 View Fig A–D, 3B–G, I–N). Location and pairs of these structures per segment differ from males to females ( Table 3 View Table 3 ), and deviations from the bilateral symmetry of their arrangement have been observed in some specimens. Up to three pairs of conspicuous laterodorsal cuticular ridges on segments 2–10 ( Figs 2 View Fig A–B, 3D–G, I–N). Cuticular hairs acicular, elongated, emerging from oval perforation sites, distributed all over the trunk cuticle. Pachycycli and ball-and-socket joints only conspicuous on segments 2–3, reduced on posterior segments ( Fig. 2 View Fig A–B, D). Apodemes on segments 7–10 ( Fig. 2 View Fig A–C). Primary pectinate fringe finely serrated; secondary pectinate fringe as a double tranverse, hairy-like, wavy row; free flaps covering the anterior part of subsequent segment ( Figs 2 View Fig A–D, 4D). Muscular scars as rounded to oval, hairless areas in laterodorsal and ventromedial positions on segments 1–10 (those of segment 1 in subdorsal and ventrolateral positions), quite inconspicuous ( Fig. 2 View Fig A–D).
Segment 1 with shortened, distally rounded middorsal process still extending beyond the posterior margin of segment ( Figs 2B View Fig , 3B View Fig , 4G). Anterolateral margins of the tergal plate as horn-shaped, short, wide, distally curved and pointed extensions ( Figs 2 View Fig A–B, D, 3A–C, H, 4A). Anterior margin of segment with a reticule-like ornamentation dorsally ( Figs 2B View Fig , 3B View Fig , 4C). Setae absent. Two pairs of sensory spots.
in ventrolateral position longitudinally arranged, and one pair in paradorsal, subdorsal and laterodorsal positions ( Figs 2 View Fig A–B, D, 3B–C, 4G). Sensory spots on this and subsequent segments as oval areas with several rows of cuticular micropapillae surrounding a single pore ( Fig. 4B, G, J View Fig ).
Segment 2 with middorsal elevation not projecting beyond the posterior margin of segment ( Figs 2B View Fig , 3D View Fig , 4E). Unpaired seta in paradorsal position, on this and following segments indifferently located to the right or to the left of the middorsal cuticular specialization, not following any particular pattern, near the anterior margin of the segment ( Figs 2B View Fig , 3D View Fig , 4E); paired setae in lateroventral position ( Figs 2 View Fig A–B, D, 3E); females furthermore with paired, sexually dimorphic setae in ventrolateral position ( Figs 2A View Fig , 3E View Fig ). Sexually dimorphic male tubes in ventromedial position ( Fig. 2D View Fig ); detailed morphology of these tubes not determined. Paired sensory spots in paradorsal, laterodorsal and ventrolateral position, the latter near the ventrolateral-ventromedial limit, not longitudinally aligned with the following ventromedial sensory spots ( Figs 2 View Fig A–B, D, 3D–E).
Segment 3 with middorsal elevation as on the preceding segment ( Figs 2B View Fig , 3F View Fig ). Unpaired seta in paradorsal position ( Figs 2B View Fig , 3F View Fig ); paired setae in laterodorsal, lateroventral and ventromedial positions ( Figs 2 View Fig A–B, 3F–G). Paired sensory spots in paradorsal, subdorsal, laterodorsal and ventromedial positions, the latter mesially shifted compared to the previous ones, aligned with those of the following segments ( Figs 2 View Fig A–B, 3F–G).
Segment 4 with middorsal elevation as on the preceding segment ( Figs 2B View Fig , 3I View Fig , 4B). Unpaired seta in paradorsal position ( Figs 2B View Fig , 3I View Fig ); paired setae in lateroventral position ( Figs 2 View Fig A–B, 3J). Paired sensory spots in paradorsal, subdorsal, laterodorsal and ventromedial positions ( Figs 2 View Fig A–B, 3I–J).
Segment 5 similar to segment 3 in the arrangement of the cuticular elevation, setae and sensory spots ( Figs 2 View Fig A–B, 4F, H).
Segment 6 similar to segment 4 in the arrangement of the cuticular elevation, setae and sensory spots ( Fig. 2 View Fig A–B).
Segment 7 similar to segments 3 and 5 in the arrangement of the cuticular elevation, setae and sensory spots ( Fig. 2 View Fig A–B).
Segment 8 with middorsal elevation as on the preceding segment ( Figs 2B View Fig , 3K View Fig ). Unpaired seta in paradorsal position ( Figs 2B View Fig , 3K View Fig ); paired setae in lateroventral position ( Fig. 2 View Fig A–B). Two pairs of sensory spots in subdorsal position, and one pair in paradorsal, laterodorsal and ventromedial positions ( Figs 2 View Fig A–B, 3K–L, 4J).
Segment 9 with middorsal process as that of segment 1 ( Figs 2B View Fig , 3M View Fig , 4I). Unpaired seta in paradorsal ( Figs 2B View Fig and 3M View Fig ); paired setae in laterodorsal and lateroventral positions ( Fig. 2 View Fig A–B). Two pairs of sensory spots in subdorsal position, and one pair in paradorsal, laterodorsal and ventromedial positions ( Figs 2 View Fig A–B, 3M–N, 4I). Nephridiopores in lateroventral position.
Segment 10 without middorsal cuticular specialization. Paired setae in lateroventral position ( Figs 2 View Fig A–C, 4I). Two pairs of sensory spots in subdorsal position, and one pair in laterodorsal and ventrolateral positions, the latter near the tergosternal junction ( Figs 2 View Fig A–C, 4I).
Segment 11 without cuticular appendages, partly retractable into segment 10 ( Fig. 4A, I View Fig ). Tergal plate triangular, with a concave and distally pointed posterior margin; sternal plates form a pair of ventral extensions rounded distally ( Figs 2 View Fig A–C, 4I). Males with two sexually dimorphic pairs of stout, thick penile spines ( Fig. 2C View Fig ). Lateral terminal spines’ length differs from males (relatively longer, LTS average ratio = 158.93 μm) to females (relatively shorter, LTS average ratio = 100.9 μm)
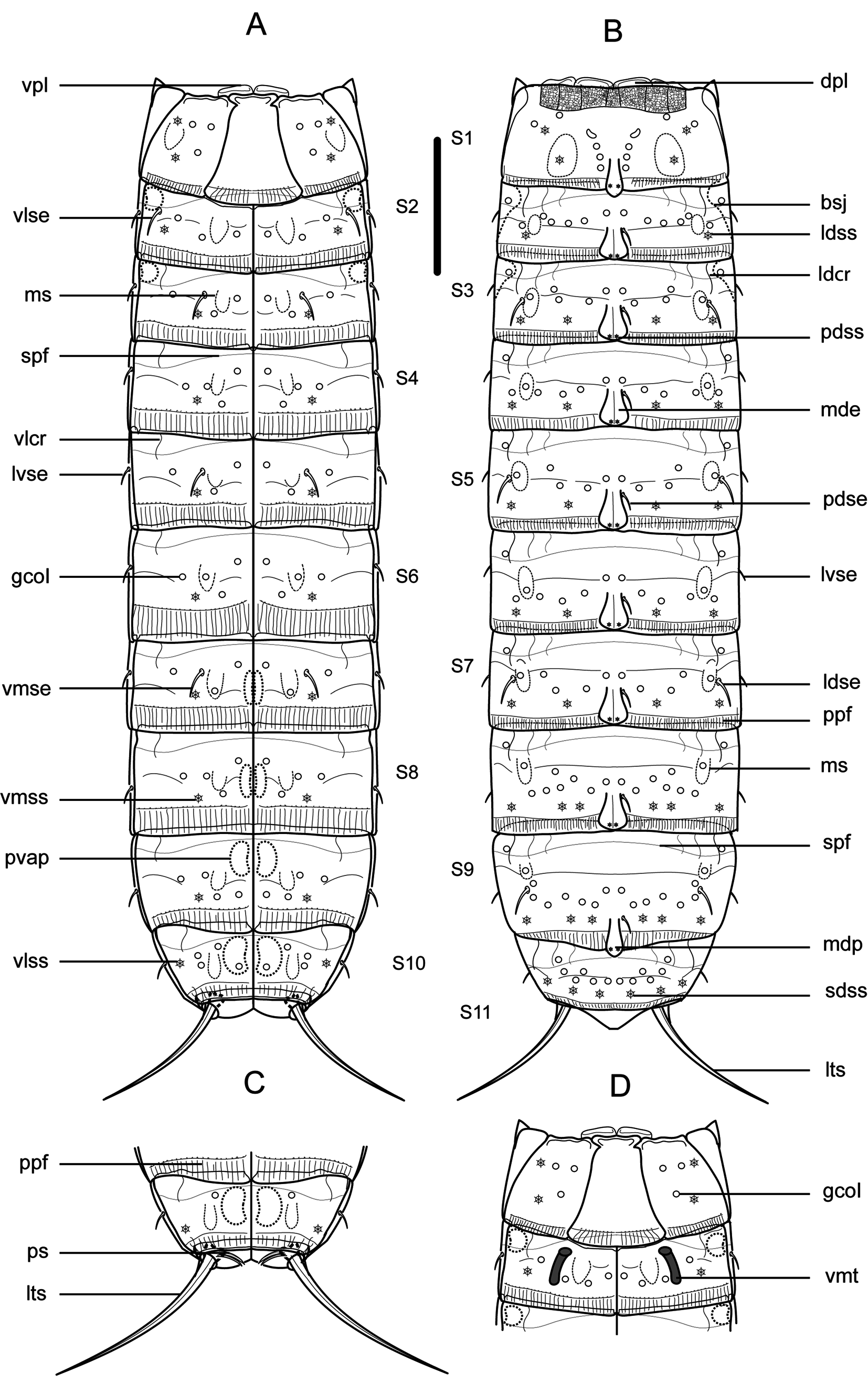
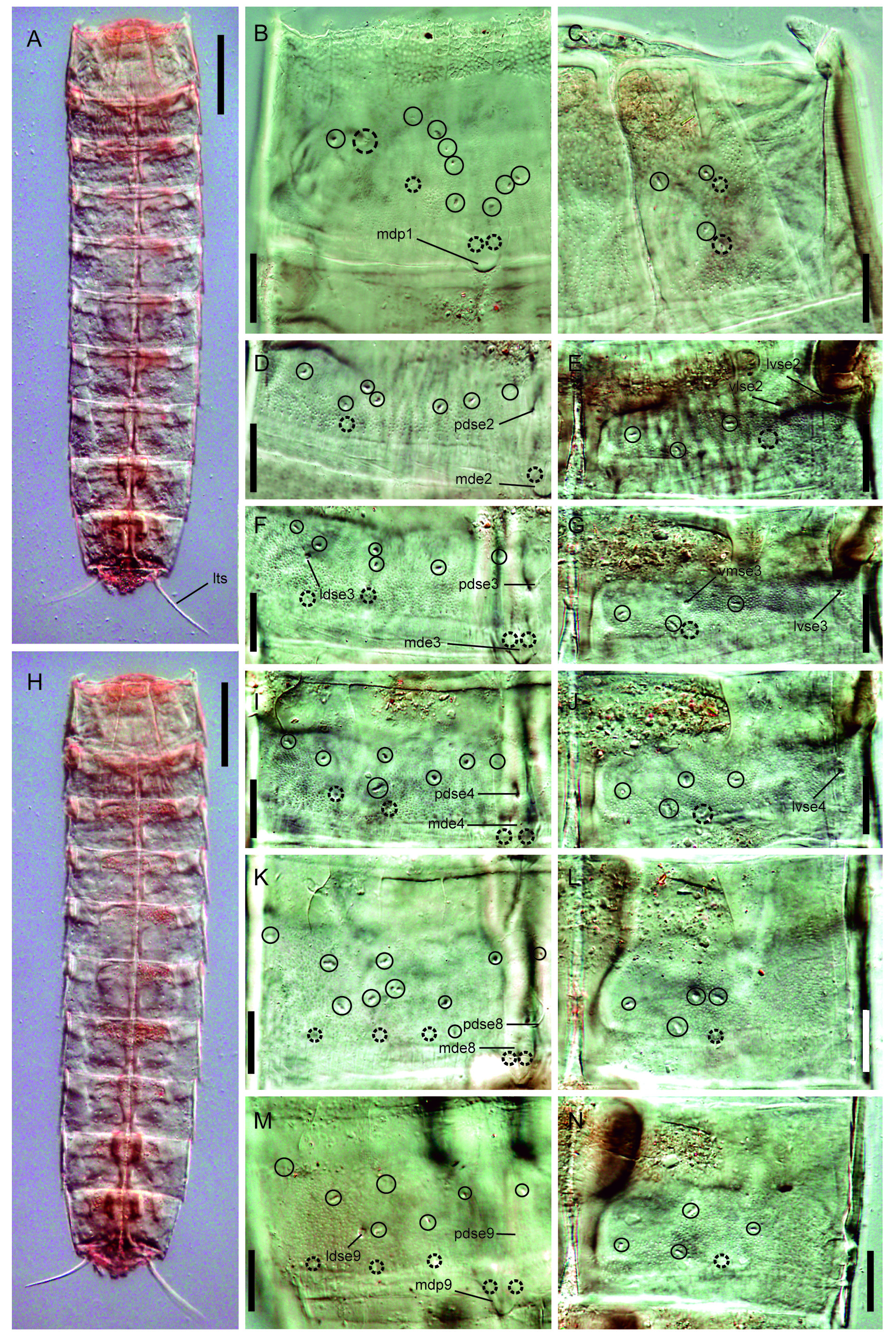
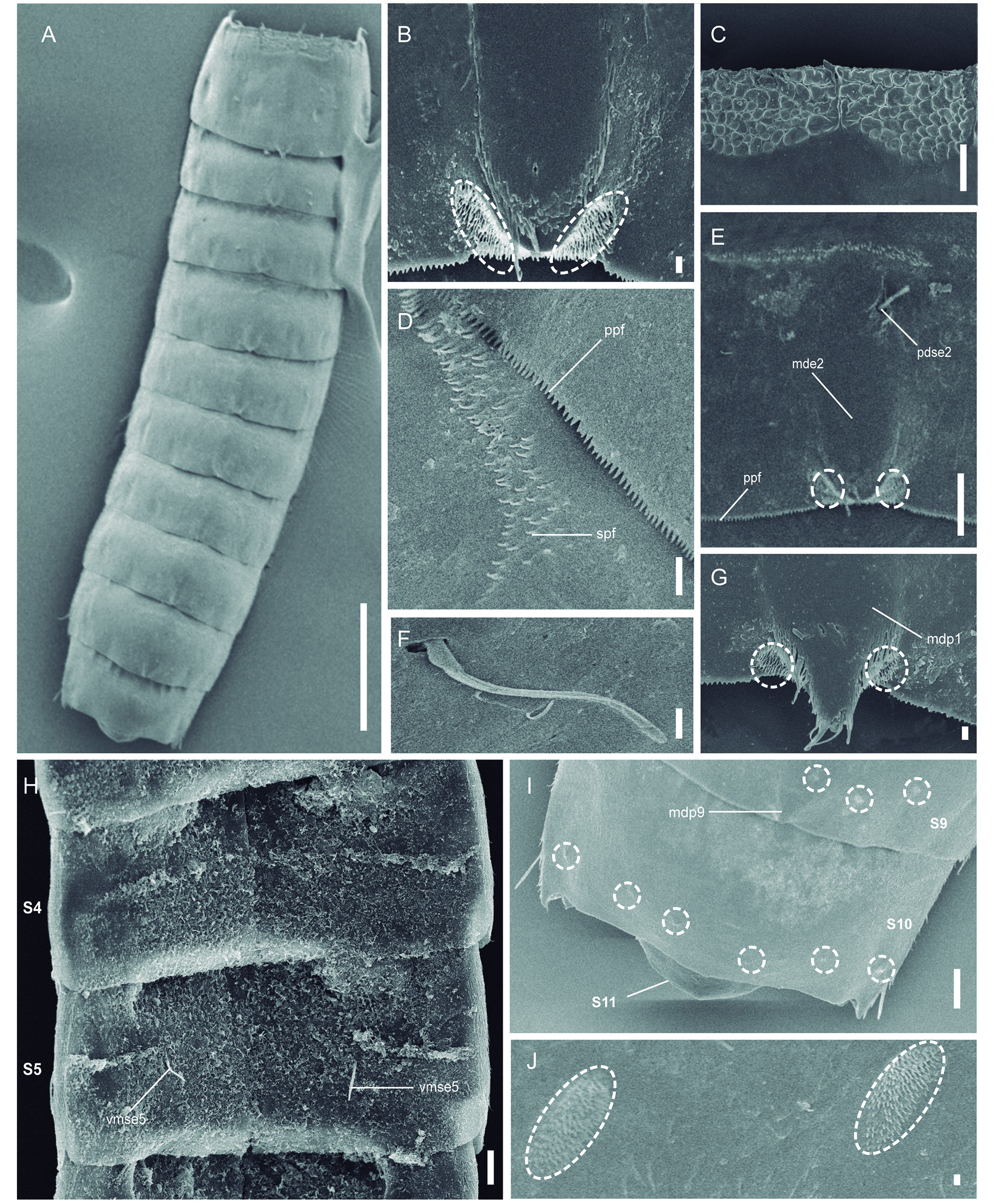
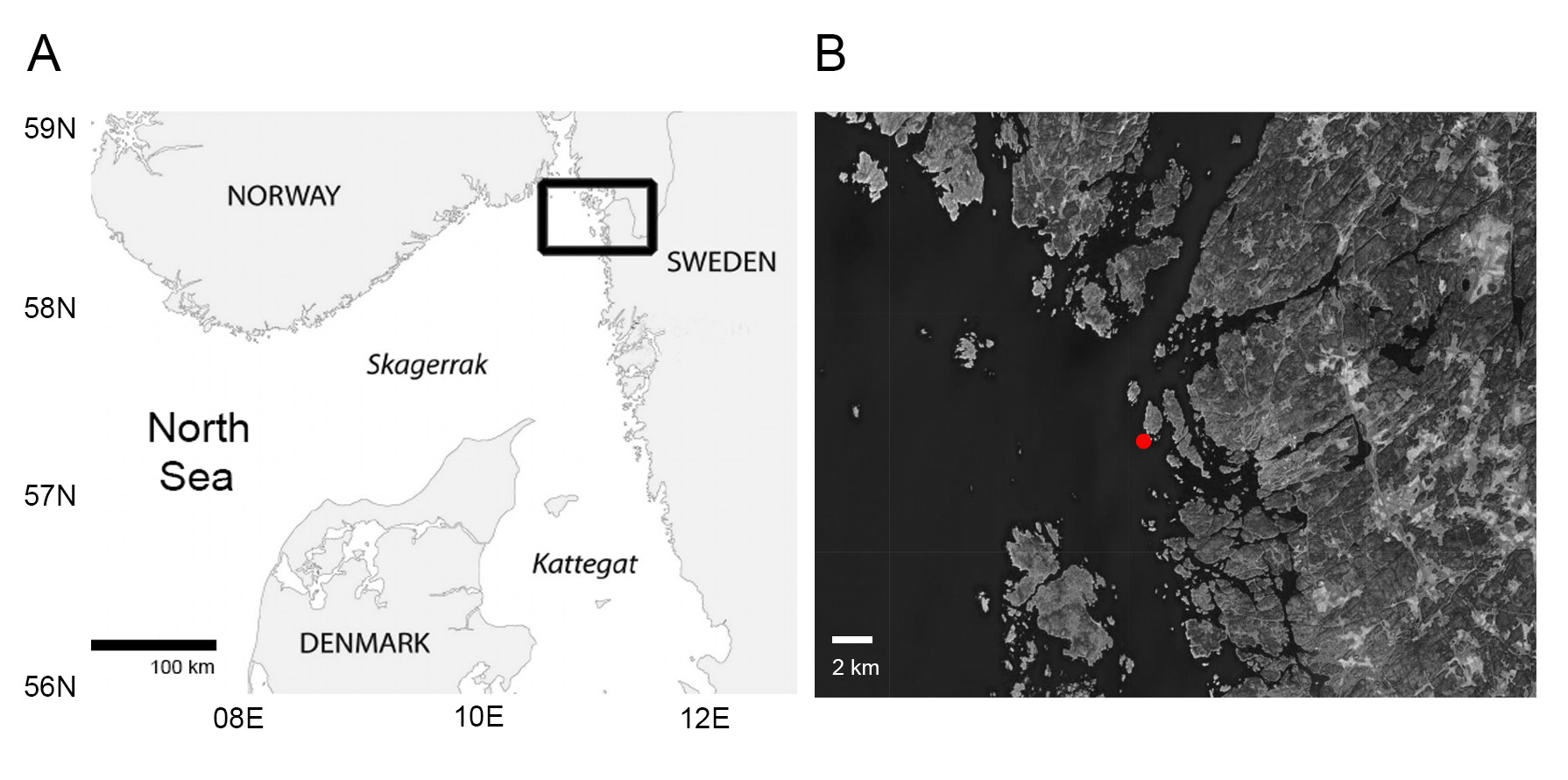
Table 1. Measurements of nine adult specimens of Setaphyes elenae sp. nov. (four males and five females) from Skagerrak. Abbreviations: LTS = lateral terminal spines; MSW = maximum sternal width (measured at segment 5); S = segments’ length (followed by number of corresponding segment); Sd = standard deviation; SSW = standard sternal width (measured at segment 10); TL = total length.
| Character | Range ♀ | Mean ♀ | Range ♂ | Mean ♂ | Total range | Total mean | Sd |
|---|---|---|---|---|---|---|---|
| TL (µm) | 612.67–722.64 | 679.192 | 614.32–647.86 | 625.51 | 612.67–722.64 | 655.33 | 44.06 |
| MSW–5 (µm) | 179.55–188.89 | 183.422 | 168.06–176.25 | 170.72 | 168.06–188.89 | 177.78 | 7.61 |
| MSW–5/TL (%) | 26–29.5 | 27 | 26.2–28.4 | 27.3 | 26–29.5 | 27.2 | 1.22 |
| SSW (µm) | 141.41–152.44 | 144.178 | 124.68–133.65 | 131.065 | 124.68–152.44 | 138.35 | 8.23 |
| SSW/TL (%) | 19.7–23 | 21.3 | 19.2–21.9 | 21 | 19.2–23 | 21.1 | 1.14 |
| S1 (µm) | 87.74–108.41 | 96.584 | 89.16–96.28 | 91.548 | 87.74–108.41 | 94.35 | 6.31 |
| S2 (µm) | 59.74–82.08 | 69.712 | 62.45–68.07 | 65.068 | 59.74–82.08 | 67.65 | 6.42 |
| S3 (µm) | 57.56–79.22 | 70.068 | 63.54–68.8 | 66.655 | 57.56–79.22 | 68.55 | 6.04 |
| S4 (µm) | 70.27–79.51 | 74.618 | 68.79–81.68 | 74.51 | 68.79–81.68 | 74.57 | 4.48 |
| S5 (µm) | 69.77–83.01 | 77.1 | 69.3–76.08 | 74.438 | 69.3–83.01 | 75.92 | 5.84 |
| S6 (µm) | 73.96–81.58 | 78.504 | 73.81–83.53 | 77.013 | 73.81–83.53 | 77.84 | 3.51 |
| S7 (µm) | 74.47–85.94 | 80.752 | 73.33–80.37 | 75.385 | 73.33–85.94 | 78.37 | 4.70 |
| S8 (µm) | 69.57–86.82 | 80.846 | 77.08–79.98 | 78.138 | 69.57–86.82 | 79.64 | 5.31 |
| S9 (µm) | 70.24–84.44 | 79.61 | 68.76–78.56 | 74.463 | 68.76–84.44 | 77.32 | 5.43 |
| S10 (µm) | 64.87–77.97 | 73.66 | 60.93–71.98 | 67.165 | 60.93–77.97 | 70.77 | 5.92 |
| S11 (µm) | 27.93–39.81 | 32.3 | 27.49–34.23 | 30.615 | 27.49–39.81 | 31.55 | 3.74 |
| LTS (µm) | 90.64–105.7 | 100.9 | 149.06–168.74 | 158.928 | 90.64–168.74 | 126.69 | 31.29 |
| LTS/TL (%) | 13.9–15.8 | 14.9 | 24–26 | 25.4 | 13.9–26 | 19.6 | 5.60 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |