
Parabuthus muelleri Prendini, 2000a: 32–38
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2003)408<0001:DOTMOP>2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5057175 |
|
persistent identifier |
https://treatment.plazi.org/id/9609F678-E97A-1A67-FF5A-F9E2FD3C627A |
|
treatment provided by |
Felipe |
|
scientific name |
Parabuthus muelleri Prendini, 2000a: 32–38 |
| status |
|
Parabuthus muelleri Prendini, 2000a: 32–38 View in CoL ,
figs. 1–9 View Fig View Figs View Figs View Figs , table 2.
Parabuthus muelleri: Prendini, 2001a: 17 View in CoL ; 2001b:
137.
TYPE MATERIAL: Holotype ♀ ( NMNW 1854 View Materials ), Namibia: Hardap Region: Maltahöhe District: Farm Onis 8, 82 km from Sesriem to Naukluft , 24°22.46̍S, 16°13.17̍E, 1260 m, 7.i.1998, L. Prendini and E. Scott . Paratype ♀ ( SAMC C4514 View Materials ), Namibia: Karas Region: Lüderitz District: Farm Plateau 38, near Aus , 26°40.62̍S, 16°31.85̍E, 1550 m, 30.xii.1997, L. Prendini and E. Scott .
DIAGNOSIS: Parabuthus muelleri falls in a group of species also including P. calvus , P. capensis , P. pallidus , and P. planicauda . This species is morphologically most easily confused with P. capensis , with which it shares the following combination of characters: metasomal segments I and II, stridulatory region extended anteriorly beyond anterodorsal edge of segment, giving a steplike appearance in lateral aspect; metasomal segments II–IV, dorsosubmedian carinae with distal spiniform granules more pronounced than preceding granules; metasomal segment IV, dorsosubmedian carinae medially discontinuous, median lateral carina continuous and distinct; metasomal segment V, dorsosubmedian carinae distinct with sharp, spiniform or subspiniform granules, and dorsolateral carinae distally obsolete. Parabuthus muelleri and P. capensis can be separated from all other Parabuthus on the basis of the following character: metasomal segment II, and to a lesser extent III, with posterodorsal edge elevated and slightly curved forward medially, forming a subtriangular Vshape.
Although morphologically similar, P. muelleri can be separated from P. capensis by several characters. The movable finger of the pedipalp chela (adult male and female) is curved ventrally in P. muelleri , such that the proximal dentate margin is distinctly emarginate when the fingers are closed (i.e., a proximal ‘‘gap’’ is evident). The emarginate condition occurs in the male of several Parabuthus species (e.g., P. granulatus , P. kalaharicus , and P. laevifrons ), but it is uncommon in female Parabuthus , and does not occur in the male or female of P. capensis . In addition, P. muelleri has a more slender metasoma, in which the median width:length percentage for metasomal segments I–V (n = 3) is 80.5% (75–86%), 78% (75–81%), 76% (72–80%), 69.5% (65–74%), and 55% (53–57%), compared with the metasoma of P. capensis , in which the median width: length percentage (n = 6) is 86% (77–95%), 73% (78–88%), 92% (79–105%), 73% (67– 79%), and 58% (53–63%). Metasomal segment III is usually broader than segments I and II in P. capensis , but this is not the case in P. muelleri . Parabuthus muelleri is further distinguished by the unusual shape of the telson, which differs from all known Parabuthus species in the presence of a distal ‘‘bulge’’ and a very short, sharply curved aculeus. The median percentage of aculeus length:telson length in P. muelleri (n = 3) is 30% (26–34%), compared with 39% (36– 42%) in P. capensis (n = 6). The two species may also usually be distinguished by the relative positions of the trichobothria on the fixed finger of the chela: eb and esb are located proximal to the basal dentate margin of the fixed finger in P. muelleri , whereas eb is located proximal to the basal dentate margin, and esb is located distal to it, in P. capensis . However, this character has been found to be polymorphic in P. capensis : in populations from the eastern part of the distributional range, esb is also located proximal to the basal dentate margin ( Prendini, 2000a).
Two characters of the newly described male can also be used to separate P. muelleri from P. capensis , notably the lobate condition of the pectinal proximal median lamella and pedipalp chelae that are not incrassate (i.e., not sexually dimorphic). The absence of sexual dimorphism of the pedipalp chelae is a particularly obvious diagnostic difference between P. muelleri and P. capensis , in which the pedipalp chelae are markedly dimorphic (the pedipalp chela manus of the male is incrassate, whereas that of the female is slender). Both characters are discussed fur ther in the description of sexual dimorphism, below.
DESCRIPTION: The following description of the male ( AMNH [ AH 3991 View Materials ]) supplements the previous descriptions ( Prendini, 2000a) of the holotype female ( NMNW 1854 View Materials ) and paratype female ( SAMC C4514 View Materials ) .
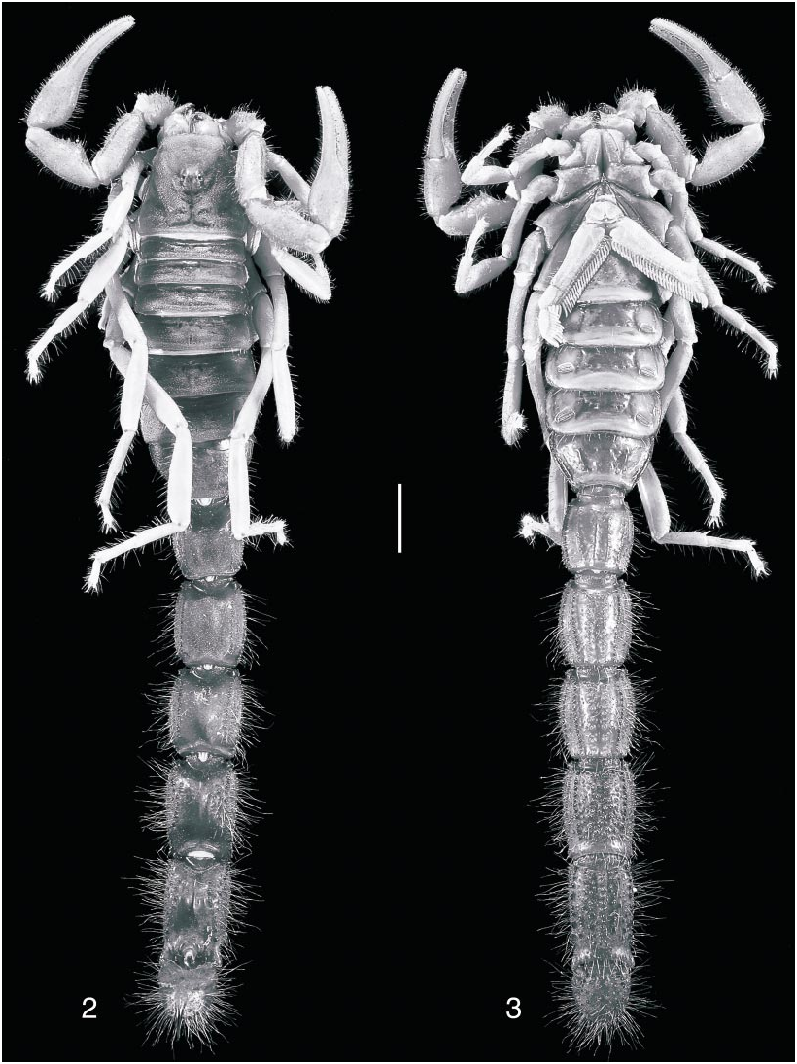
Color: Carapace, chelicerae, tergites, sternites, and metasomal segments I–III: Cinnamon no. 123A. Metasomal segments IV, V and telson: Burnt Sienna no. 132. Pedipalps and legs: Clay Color no. 123B. Pectines: Chamois no. 123D. Metasomal segments IV, V and telson are distinctly darker than segments I–III, whereas pedipalps and legs are distinctly paler than carapace, mesosoma, metasoma, and telson ( figs. 2, 3 View Figs ).
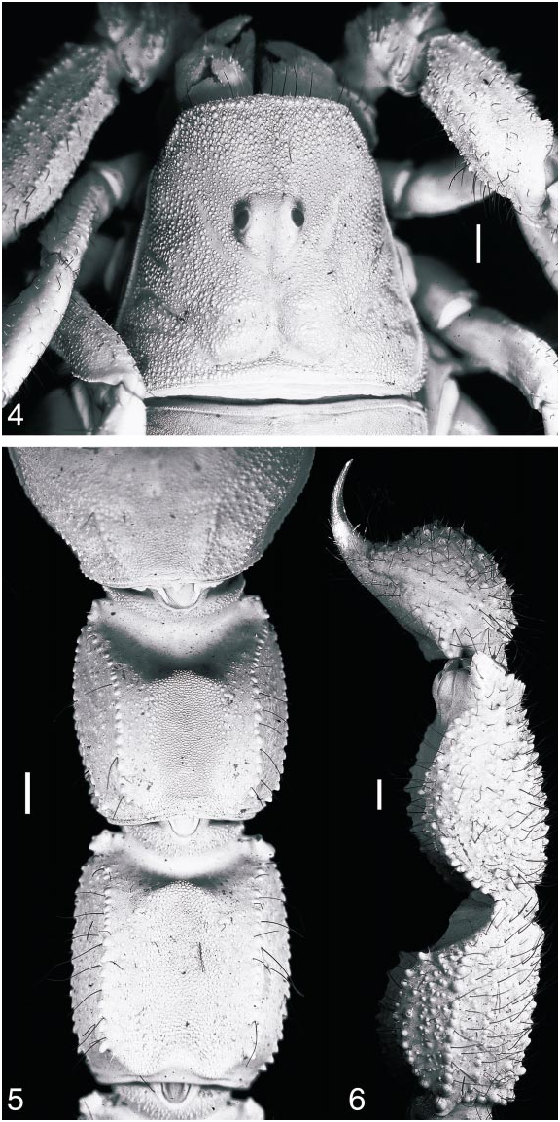
Carapace: Carapace with sulci, without carinae, and covered entirely by uniform, coarse granulation, becoming coarser on interocular and posterolateral surfaces. Anteri or margin of carapace procurved; posterior margin straight. Five pairs of lateral ocelli. Median ocelli considerably larger than lateral ocelli, situated anteromedially ( fig. 4 View Figs ). Ocular tubercle with pair of smooth superciliary carinae, protruding slightly above median ocelli. Anteromedian furrow shallow; posteromedian furrow shallow anteriorly, becoming deeper posteriorly; posterolateral furrows shallow, wide, curved; posteromarginal furrow narrow, deep.
Chelicerae: Movable finger with distal external and distal internal teeth equal, opposable. Ventral aspect of fingers and manus with long, dense macrosetae. Fixed finger with a pair of denticles on the ventral surface.
Sternum: Subtriangular ( fig. 3 View Figs ). Median longitudinal furrow Yshaped, shallow anteriorly, deep and narrow posteriorly.
TABLE 3 Length (steps), Fit (f i), Consistency Indices (CIs), and Retention Indices (RIs) of 50 Informative Characters Scored Among 25 Species of the Genus Parabuthus Pocock, 1890 Final weights obtained with succesive weighting (SW) are also reported.
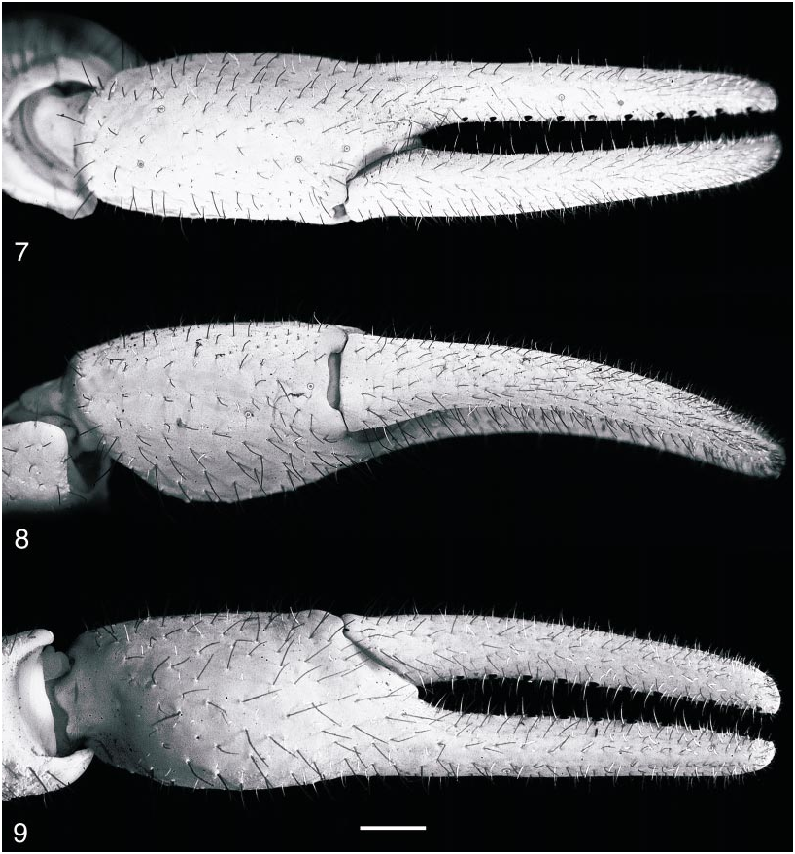
Pedipalps: Pedipalps covered in short macrosetae ( figs. 7–9 View Figs ). Femur finely and uniformly granular; pentacarinate, all carinae distinct, granular, except for internomedian carina, comprising spiniform granules. Patella finely and uniformly granular; carinae absent or obsolete; dorsointernal and ventrointernal carinae each comprising a row of granules proximally; internomedian carina comprising a large spiniform granule, proximally, and a few smaller granules, distally. Chela smooth; carinae absent. Chela short, slender, length along ventroexternal carina 28% greater than chela width and 30% greater than chela height; length of movable finger 45% greater than length along ventroexternal carina. Chela fixed finger slightly curved dorsally and movable finger slightly curved ventrally, such that proximal dentate margin emarginate when fingers are closed ( fig. 7 View Figs ). Dentate margins of chela fingers each with 11 oblique granular rows, each comprising 4–6 small granules and a large proximal granule, flanked by an inner and an outer accessory granule; chela fingers each with a terminal denticle.
Trichobothria: Orthobothriotaxic, type A, α configuration, with the following segment totals ( figs. 7–12 View Figs View Figs ): femur, 11 (5 dorsal, 4 internal, 2 external), patella, 13 (5 dorsal, 1 internal, 7 external), and chela, 15 (8 manus, 7 fixed finger). Total number of trichobothria per pedipalp, 39. Chela with eb located proximal to basal dentate margin of fixed finger and esb located just distal; dt almost level with et; db equidistant between est and esb. Patella with esb 2 slightly distal to esb 1. Femur with d 2 on proximointernal side of dorsointernal carina; d 3 distal to d 2; d 4 equidistant between d 3 and d 5.
Mesosoma: Pretergites smooth and shiny, granular along posterior margins. Posttergites entirely coarsely granular, granulation becoming coarser distally ( fig. 2 View Figs ); I–VII each with a weakly developed, granular median
3. Ventral aspect. Scale bar = 5 mm.
carina; VII additionally with distinct pairs of costate granular dorsosubmedian and dorsolateral carinae, and with welldeveloped stridulatory region between dorsosubmedian carinae, consisting of round to slightly crescentshaped granules reaching the posterior margin. Sternites entirely smooth, except for posterolateral surfaces of sternite VII, which are sparsely granular; lateral and distal margins each with a row of sparsely distributed macrosetae; sternite VII with weakly developed pairs of costate ventrosubmedian and ventrolateral carinae.
Pectines: First proximal median lamella of each pecten suboval, mesially enlarged and lobate ( fig. 3 View Figs ). Pectinal teeth: 36/36.
Genital operculum: Completely divided longitudinally. Genital papillae present.
Legs: Tibia III and IV with spurs; retrolateral margins with scattered macrosetae. Basitarsi I and II only slightly compressed dorsoventrally, retrolateral margins each with dense row of long, fine macrosetae. Telotarsi each with paired ventrosubmedian rows of fine macrosetae. Telotarsal laterodistal lobes truncated; median dorsal lobes extending to ungues. Telotarsal ungues short, distinctly curved, and equal in length.
Metasoma and telson: Metasomal segments I–V width/length ratio progressively decreasing, width percentage of length 86% for I, 81% for II, 80% for III, 68% for IV, and 53% for V (table 4). Telson oval, globose ( fig. 6 View Figs ), height 66% of length, with flattened dorsal surface and rounded ventral surface; vesicle not significantly narrower than metasomal segment V, width 90% of metasomal segment V. Metasoma entirely granular, except for ventromedian surfaces of segments I and II, and dorsomedian surfaces of segments IV, V and telson. Metasomal segments I and II each with a welldeveloped stridulatory region on the dorsomedian surface, consisting of round to slightly crescentshaped fine granules extending to the posterior margin ( fig. 5 View Figs ); stridulatory region of segment III narrow, virtually obsolete, consisting of a few granules in the proximal third of the segment; segment II, and to a lesser extent III, with posterodorsal edge elevated and slightly curved forward medially, forming a subtriangular Vshape ( fig. 5 View Figs ). Metasoma densely covered with long macrosetae, especially on the ventral surface of the telson. Metasomal segments I–IV each with 10 carinae, but segment IV with ventrosubmedian and median lateral carinae distinct only in the proximal half of the segment; segment V with seven carinae, including a single, obsolete granular ventromedian carina, a pair of distinct ventrolateral carinae, a pair of dorsolateral carinae, distinct only in the proximal half of the segment, and a pair of dorsosubmedian carinae reduced to a few prominent rounded or subspiniform granules medially. Metasomal segments I–V with dorsosubmedian carinae converging distally in segment I, subparallel in segments II–V; ventrolateral carinae converging distally in segments I–III, subparallel in segment IV, diverging in segment V. All metasomal carinae costate granular to granular, except for ventrosubmedian and ventrolateral carinae of segment I, which are costate to costate granular. Metasomal segments II–IV with distal granules of dorsosubmedian carinae very slightly enlarged, rounded; segments II and III with distal granules of ventrosubmedian carinae and, to much a lesser extent, ventrolateral carinae, enlarged, obtuse, and elevat ed; segment V with subdistal granules of ventrolateral carinae enlarged into laterally compressed, lobate processes. Telson with a distal ‘‘bulge’’ and a very short, sharply curved aculeus ( fig. 6 View Figs ); aculeus length percentage of vesicle length, 35%.
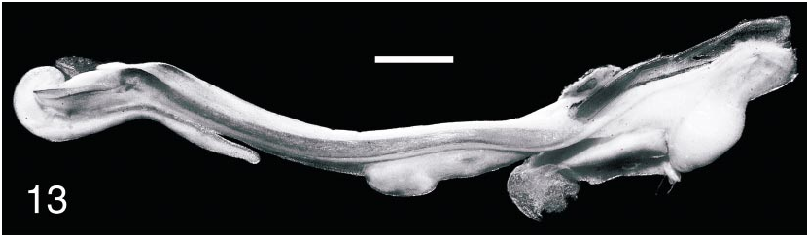
Hemispermatophore: A hemispermatophore was dissected from the male, confirming that it is adult. The hemispermatophore is typical of other species of Parabuthus , characteristically flagelliform, with pars recta parallel to axis of distal lamina ( fig. 13 View Fig ).
Sexual dimorphism: Unlike most species of Parabuthus , in which the pedipalp chela manus of the adult male is noticeably incrassate (bulbous or swollen), compared with the more slender manus of the adult female ( Pocock, 1889, 1890, 1902; Kraepelin, 1899, 1908; Purcell, 1898, 1899, 1901; Werner, 1916; Hewitt, 1913, 1915, 1918; Eastwood, 1977; Lamoral, 1977, 1979, 1980; Newlands and Martindale, 1980; Prendini, 2001a), the adult male of P. muelleri is not sexually dimorphic in this respect. The chela width: length and height:length ratios of 72% and 70%, respectively, in the male do not differ significantly from the equivalent ratios of 74% and 69%, respectively, in the female. Parabuthus calvus , P. nanus , P. pallidus , and P. planicauda are the only other species of the genus in which the pedipalp chelae of the adult male are not sexually dimorphic. The absence of sexual dimorphism of the pedipalp chelae provides an additional obvious diagnostic character for separating P. muelleri from the closely related and morphologically similar P. capensis . The male of P. muelleri also displays weakly lobate prox imal median lamellae of the pectines, a character observed in the female of most Parabuthus species— P. granulatus and P. kalaharicus are exceptions—but otherwise only in the male of P. calvus and P. planicauda ( Purcell, 1898, 1901; Pocock, 1902; Kraepelin, 1908; Hewitt, 1918; Werner, 1934; Lawrence, 1955; Eastwood, 1977; Lamoral, 1979; Sissom, 1994; Prendini, 2001a). In other respects, the differences between the
TABLE 4 Meristic Data for Holotype Female (NMNW 1854), Paratype Female (SAMC C4514), and Newly Discovered Male (AMNH [AH 3991]) of Parabuthus muelleri Prendini, 2000 Male Parabuthus capensis (Ehrenberg, 1831) from Vanrhynsdorp, Western Cape Province, South Africa (SAMC C4565) included for comparison. Measurements follow Stahnke (1970), Lamoral (1979), and Prendini (2000a).
male and female of P. muelleri are typical of other species of Parabuthus . The male exhibits genital papillae, a greater pectinal tooth count, increased granulation and setation (particularly of the pedipalps, metasoma, and telson), and is proportionally more slen der than the female.
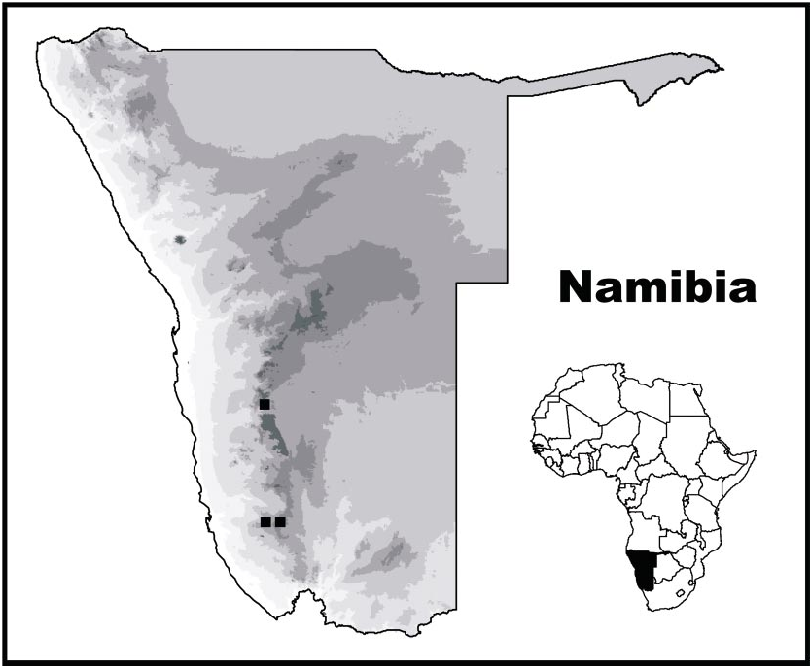
DISTRIBUTION: Parabuthus muelleri is endemic to Namibia ( fig. 14 View Fig ) and presently recorded from two neighboring localities in the Lüderitz District (Karas Region) and a third locality, ca. 260 km north, in the Maltahöhe District (Hardap Region). Despite the large distance between the first two localities and the third, the habitat in which the specimens were collected (at the two localities for which habitat data are available) was remarkably similar ( Prendini, 2000a). The specimens were each found in an area of level ground, with chalky soil, calcrete nodes, and dolomitic rocks. The localities at which they were collected both occur along an extensive ridge of dolomite, extending from the HuibHoch Plateau in the south (Plateau is situated at the northern end) to the Naukluft mountains in the north (Onis is ca. 20 km south of the Naukluft). Thus, the two localities are fairly close to the northern and southern edges of this geological formation. It is not known whether P. muelleri is restricted to this formation, but the occurrence of two of the three known specimens in such similar, albeit distantly located habitats suggests that the species may occur in similar habitats throughout the intervening areas ( Prendini, 2000a).
ECOLOGY: The holotype and paratype of P. muelleri were collected in a region of com pacted, chalky soil, calcrete nodes, and dolomite rocks. The holotype was excavated from the burrow of a scorpionid, Opistophthalmus opinatus (Simon, 1888) , which it had preyed on, and was also syntopic with O. scabrifrons Hewitt, 1918 . The paratype was found sitting motionless on a stone at night and was syntopic with the ischnurid Hadogenes tityrus (Simon, 1888) , the scorpionids Opistophthalmus gigas Purcell, 1898 and O. scabrifrons , and also with Parabuthus granulatus and P. villosus . According to A. Harington’s collecting notes, the newly described male specimen was collected in an area with P. villosus and P. laevifrons , with the latter probably being a misidentification of P. stridulus .
ADDITIONAL MATERIAL EXAMINED: Ƌ (AMNH [AH 3991]), Namibia: Karas Region: Lüderitz District: locality uncertain, probably Aus [26°41̍S, 16°15̍E], A. Harington. Unfortunately, the collection locality data for this specimen are ambiguous. Harington suggests two possible localities, the other being Wortel (Khomas Region: Windhoek District: 23°08̍S, 17°10̍E). Aus is clearly the more plausible of the two alternatives, given its close proximity to Plateau, the collection locality of the paratype female. Indeed, it is quite possible that the male specimen also originated from Plateau, where Harington collected extensively and also received material donated by the owners of the property, H. and W. Erni. The possibility that the male specimen was collected at Plateau also seems more likely in view of the considerably different habitats at Aus and Plateau, which have few scorpion species in common, de spite their proximity. The Aus Mountains are part of a granitic formation completely separated from the dolomitic HuibHoch Plateau by a sandy plain approximately 10 km across at the narrowest point. No scorpions associated with rocky habitats in either the Aus Mountains or the HuibHoch Plateau have been collected on this plain, which probably represents a significant barrier to their dispersal, as has been demonstrated elsewhere for lithophilous and lapidicolous scorpion species ( Newlands, 1972; Prendini, 2001b).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parabuthus muelleri Prendini, 2000a: 32–38
| PRENDINI, LORENZO 2003 |
Parabuthus muelleri: Prendini, 2001a: 17
| Prendini, L. 2001: 17 |
Parabuthus muelleri
| Prendini, L. 2000: 38 |