
Anurida abashiriensis, Babenko & Nakamori, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4425.3.10 |
|
publication LSID |
lsid:zoobank.org:pub:EB11C783-6766-4C40-AEB6-F95F7E2C2162 |
|
DOI |
https://doi.org/10.5281/zenodo.5968659 |
|
persistent identifier |
https://treatment.plazi.org/id/984587CD-FF88-FF9B-FF1E-FA7B7A8FC5B9 |
|
treatment provided by |
Plazi |
|
scientific name |
Anurida abashiriensis |
| status |
sp. nov. |
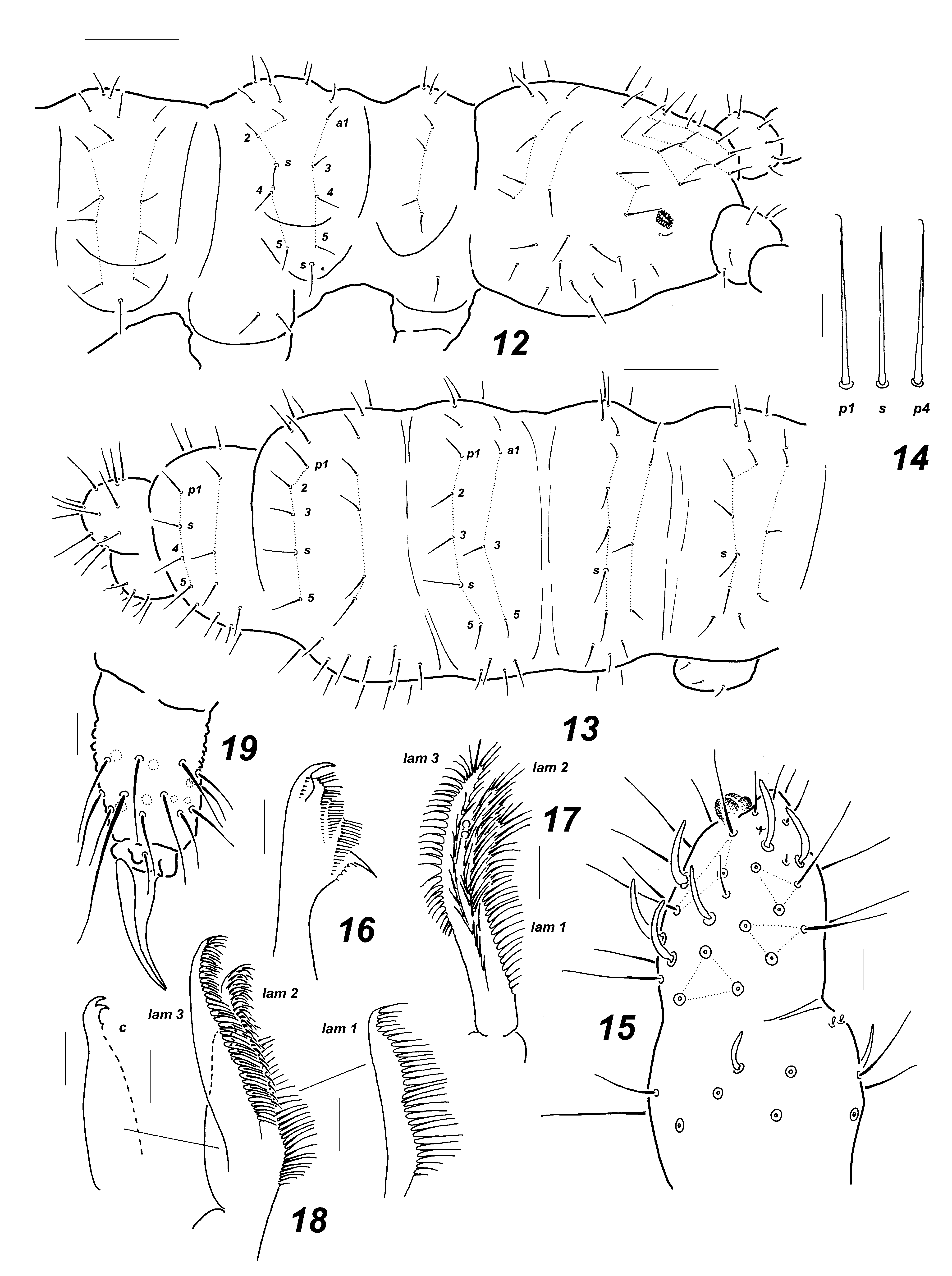
Anurida abashiriensis View in CoL sp. nov. ( Figs 12–19 View FIGURES 12–19 )
Diagnosis. Blind species of genus Anurida characterized by presence of six blunt sensilla on Ant.4 (S1–S4, S8–S9), oval PAO with about 15–17 lobes, unique mandibles with three intermediate rows of long thin filaments between apical and basal teeth, elongate maxilla with all three lamellae reaching well beyond tip of capitulum, Th.2–3 with p2-seta set posterior to p1 and complete absence of m-setae on all terga.
Type material. Holotype (male) on slide, Japan, Hokkaido Island, seashore E of Abashiri [ N 43.9942°, E 144.2961°], sandy/shingly beach (floatation), 18 viii 2016, M. Potapov & N. Kuznetsova leg. ( MSPU) GoogleMaps . Paratypes, 3 females, 6 males and 6 juveniles, same data as holotype ( MSPU) GoogleMaps ; 1 female and 3 males, same locality as holotype, 31 viii 2017, T. Nakamori leg. ( TRPM) GoogleMaps
Description. Length without antennae 1.3–1.7 mm, holotype—1.62 mm. Colour very bright with scarce grey pigment granules all over body. Body shape typical of genus, rather slender and elongate, Abd.6 rounded, not truncate. Integument granulation uniform and rather coarse.
Ocelli absent. PAO elliptical consisting of about 15–17 lobes. Antennae typical of genus, conical and clearly shorter than head. Ant.4 with 3-lobed apical bulb, six usual curved sensilla, subapical ms and organite present ( Fig. 15 View FIGURES 12–19 ). AO on Ant.3 typical, outer sensilla widely separated, small ms present ventrally. Ant.1–2 with 7 and 12 setae, respectively. Labral formula as 4/2-3-5-2, lateral pair of prelabral setae widely separated. Labium with 11 setae on each side including three setae on its apical part also armed with two small sensillar papillae. Mandibles very characteristic ( Fig. 16 View FIGURES 12–19 ) with strong apical and basal teeth and three intermediate oblique, transverse rows of long thin filaments. Maxillary capitulum with at least two small apical teeth on main part (hardly detectable) being covered by three lamellae with very long marginal filaments, all lamellae reaching well beyond tip of capitulum ( Figs 17–18 View FIGURES 12–19 ).
Dorsal setae weakly differentiated, longer on abdominal tip, sensilla hardly distinguishable from ordinary setae ( Fig. 14 View FIGURES 12–19 ). Lateral microsensilla (ms) present only on Th.2. Main characteristics of dorsal chaetotaxy: tergum of Th.1 with at least 4 +4 setae, sometimes up to 6 setae on each side. Th.2–3 with setae p 2 set posterior to p1 and rather close to midline ( Fig. 12 View FIGURES 12–19 ) given effect of 3+3 axial setae, m-row almost completely reduced on thorax (only lateral sensilla m6 present) and abdomen. Similar tendency of backward position of setae p2 also characteristic of some abdominal terga, especially of Abd.4 ( Fig. 13 View FIGURES 12–19 ). Thoracic sterna without setae. Ventral tube with 3+3 distal setae. Furca remnant as narrow, longitudinal area without secondary granulation in midsection of Abd.4 sternum.
Legs 1–3 chaetotaxy as following: upper subcoxae—1, 2, 2; lower subcoxae—0, 2, 2; coxae—3, 6(7), (7)8; trochanters—6, 6, 6; femora—13, 12(13), 11; tibiotarsi—19, 19, 18 setae, respectively. Tibiotarsal seta A1 longer than other setae of A-whorl, pointed ( Fig. 19 View FIGURES 12–19 ). Roundish cuticular papillae on outer side of subcoxae developed but rather small. Unguis without inner or lateral tooth.
DNA barcoding. DNA sequences of partial regions of mitochondrial cytochrome c oxidase subunit I gene, 16S ribosomal RNA gene, and nuclear 28S ribosomal RNA gene were determined as described in Potapov et al. (2017). The sequences obtained from two paratypes (TRPM-AAr-0000752 (male) and TRPM-AAr-0000754 (female), same locality as holotype, 31 viii 2017, T. Nakamori leg.) were submitted to GenBank under accession nos. LC372827 View Materials – LC372832 View Materials .
Etymology. Named after Abashiri, a city in close proximity to the type locality.
Affinities. A. abashiriensis sp. nov. is a very distinctive species and it can hardly be compared with any related collembolan species. Thus, maxillae with three serrated lamellae are highly common for the Anurida complex but only few known species, namely Anurida dentata Christiansen & Bellinger, 1980 [Midwestern US], A. kaya Babenko, 1998 [the Caucasus] and Gastranurida denisi ( Bagnall, 1939) [Europe], possess elongate maxillary head with long, ciliated/ serrated lamellae, all passing well beyond its capitulum. None of these species is blind and has mandibles comparable with those of the new species. The second main diagnostic character of the new species is a unique shape of its mandibles, which however might be derived from both “dentate” mandibles typical of the genus Anurida and “special” (both these term were used by Jordana et al. 2012) ones of Gas tranurida Bagnall, 1949 or Anuridella Willem, 1906 .
The only known species of the former monotypic genus can be easily distinguished from A. abashiriensis sp. nov. due to pronounced plurichaetosis and strong differentiation of dorsal setae. All members of Anuridella have unusual slender body shape and peculiar thoracic chaetotaxy with p 1 in forward position and p2 shifted closer to mid-line ( Fjellberg 1998). Both traits are also typical of A. abashiriensis sp. nov. However, contrary to representatives of Anuridella , the new species has a complete tibiotarsal chaetotaxy, much longer maxillary lamellae and only traces of integumental coxal lobes typical of many Anuridella .
Taxonomic remarks. A generic position of the new species is obscure. A number of Pseudachorutinae genera, viz. Anurida Laboulbène, 1865 , Aphoromma MacGillivray, 1893 , Anuridella Willem, 1906 , and Gastranurida Bagnall, 1949 , characterized by the PAO presence and the absence of seta L on labium, developed mandible, maxillae with several serrated lamella and almost complete reduction of furca, have been designated. Two former genera, i.e. Anurida and Aphoromma , differing only in presence/absence of ocelli ( Bagnall 1939) were not acknowledged by most taxonomists ( Stach 1949; Gisin 1960; Massoud 1967; Christiansen & Bellinger 1980; Fjellberg 1998; Bellinger et al. 1996–2017), more rarely treated as subgenera ( Yosii 1977; Kuznetsova 1988). Anuridella and Gastranurida are listed on www. collembola .org as separated genera although independence of Anuridella and especially Gastranurida from Anurida is also a subject of long discussion. Thus, Gisin (1960) considered all of them as synonyms; Massoud (1967) accepted all three genera as valid; Christiansen & Bellinger (1980) used the name Anuridella as a subgenus of Anurida , whereas Lawrence & Goto (1968) and Fjellberg (1998) treated Anuridella as a separated genus but considered G. gisini , the type species of Gastranurida , as a member of the genus Anurida .
Both Anuridella and Gastranurida include littoral inhabitants characterized by hypertrophy of mouthparts (mandible and/or maxillae) and appear to be rather similar in main structural features. Among principal diagnostic characters of the world genera of Pseudachorutinae listed by Jordana et al. (2012, p. 53, table 2) there is only a single distinction—three ocelli in the only known species of Gastranurida and none in Anuridella , the character which displays much intrageneric variation throughout the collembolan system. Massoud (1967) asserted that the genus Gastranurida differs from Anurida by two unique characters: mandibles with numerous serrate teeth (“dents nombreuses et crénelées”) and the presence of specific structure on sterna of Abd.2 not known in other genera of Collembola (“de bourrelet sur le deuxième segment abdominal, n’ayant son équivalent chez aucun genre de Collembole”). In fact, the latter is just a furca remnant situated near the border between Abd.3 and Abd.4. Existed differences in mandible shape between all three genera appear to be adaptive and may not reflect close relationship. In general, we believe that recent generic system
within the Anurida complex of Pseudachorutinae is far from ideal and therefore prefer to treat describing intermediate form as a member of admittedly heterogenic genus Anurida .
Distribution. Known only from the type-locality.
| TRPM |
Tottori Prefectural Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anurida abashiriensis
| Babenko, Anatoly & Nakamori, Taizo 2018 |
Anurida abashiriensis
| Babenko & Nakamori 2018 |
Gastranurida
| Bagnall 1949 |
Gastranurida
| Bagnall 1949 |
Gastranurida
| Bagnall 1949 |
Anuridella
| Willem 1906 |
Pseudachorutinae
| Börner 1906 |
Anuridella
| Willem 1906 |
Pseudachorutinae
| Börner 1906 |
Anurida
| Laboulbene 1865 |
Anurida
| Laboulbene 1865 |
Anurida
| Laboulbene 1865 |