
Tanaissus microthymus Bird & Bamber
|
publication ID |
https://doi.org/ 10.5281/zenodo.187828 |
|
DOI |
https://doi.org/10.5281/zenodo.6223259 |
|
persistent identifier |
https://treatment.plazi.org/id/992487CD-7B7B-FFD6-E2B1-FF37FBFDD69F |
|
treatment provided by |
Plazi |
|
scientific name |
Tanaissus microthymus Bird & Bamber |
| status |
sp. nov. |
Tanaissus microthymus Bird & Bamber View in CoL sp. nov.
Figures 7–11 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11
Synonymy:? T. lilljeborgi Makkaveeva, 1971 , non- Leptognathia lilljeborgi Stebbing, 1891
Material examined. Holotype non-ovigerous female (NHM.2008.4969), 1.76 mm. Station ORI 2, off the northern coast of Israel, 32º 28.472 N 034º 52.316 E, 15.8 m depth, 26 May 2005. Allotype: terminal (‘swimming’) male (NHM.2008.4970), 2.83 mm; same sample as holotype. Paratypes: 249 specimens ( TAU AR 28451), same sample data as holotype; 161 specimens including 3 males (NHM.2008.4913–4922), ORI 1, 32°27.904’N 034°52.190’E, 15.8 m depth, 26 May 2005; 65 specimens ( TAU AR 28452), ORI 3, 32°27.362’N 0 34°52. 0 28’E, 16.2 m depth, 26 May 2005. 57 specimens, including 4 males (NHM.2008.4923–4932), Station AGAN 6, 31º51.342’N 034º39.050’E, 11.8 m depth; 17 females, 1 male (NHM.2008.4933–4942), Station AGAN 7, 31º51.446’N 034º39.453’E, 7.4 m depth; 47 specimens, including 2 males ( TAU AR 28453), Station AGAN 11, 31º51.014’N 034º38.992’E, 10.1 m depth; all 0 2 May 2007; 12 females, 2 males, 8 juveniles ( TAU AR 28450), MI 5, 31°51.237’N 034°39.301’E, 10 m depth, 2 May 2007.
Other material: 1 specimen, ORI 13, 32°27.510’N 034°52.316’E, 5.5 m depth, 12 July 2005; all clean fine sand. 1 manca-II, two mancae-III, four juveniles, 68 neuters/non-ovigerous/post-ovigerous females (one dissected on microslide), 36 ovigerous females, eight males, same sample data as holotype. 1 specimen, MI 2, 31°50.955’N 034°39.347’E, 5.0 m depth, 19 September 2006; 23 specimens, MI 5, 31°51.237’N 034°39.301’E, 10 m depth, 17 September 2006. 35 specimens, MI 3, 31°51.472'N 034°39.553'E, 4.7 m depth; 24 specimens, MI 4, 31°51.733’N 034°39.699’E, 4.4 m depth; 126 specimens, MI 5; all 20 October 2007. all coarse sand with shell. 34 specimens, HMA DA 331°50.812’N 034°39.161’E, 6.4 m depth; 2 specimens, HMA DA22, 31° 51.612’N 034° 39.187’E, 12 m depth; 8 specimens, HMA DA23, 31° 51.309’N 034° 39.303’E, 8.7 m depth; 40 specimens, HMA DA60, 31° 51.031’N 034° 39.247’E, 7.3 m depth; 61 specimens, HMA DA62, 31° 51.731’N 034° 39.568’E, 7.6 m depth; 8 specimens, HMA DA63, 31° 51.797’N 034° 39.068’E, 14 m depth; all 20 October 2007.
6182 specimens, NMI stations (see Appendix 2A). 1313 further specimens, AGAN stations (see Appendix 2B).
Diagnosis: typical Tanaissus , female with pleon about as long as pereonites 5 and 6 combined; mandible molar acuminate, single-pointed; cheliped propodus with bifid distal crest; fixed finger with one distal tooth on incisive margin. Male with very elongate cephalothorax, pereonites 2–5 about as long as broad, antennule seven-articled; cheliped carpus with finger-like ventral process and small distal shield.
Description of non-ovigerous female (neuter). Body ( Figure 7 View FIGURE 7 A) slender, 8.7 times as long as wide, 1.76 mm long. (Other paratype neuters/non-ovigerous females, length 1.2–1.7 mm). Cephalothorax as long as pereonites 1–3 combined, rostral half much narrower than posterior, with rounded, crenulate rostral margin ( Figure 7 View FIGURE 7 B) and two dorsal crenulate ridges. Pereonites all more or less rectilinear, pereonite-1 shortest, pereonites 1–6 0.4, 0.78, 0.92, 1.04, 0.86 and 0.71 times as long as broad respectively. Pleon just longer than pereonites 5–6 combined; pleonite-5 longest; shallow epimera with single seta. Pleotelson ( Figure 7 View FIGURE 7 C) as long as two preceding pleonites, weakly tapering and with shallow convex posterior margin bearing four setae (two penicillate).
Antennule ( Figure 7 View FIGURE 7 D) slender, 0.85 times as long as cephalothorax, three-articled, although articulation of fourth small terminal article (flagellum) is suggested; peduncle article-1 longer than articles 2 and 3 combined, with proximal and distal clusters of three penicillate setae and single simple seta; article-2 longer than broad with one penicillate and two simple distal setae; article-3 three times as long as broad, with about five simple terminal setae and one penicillate seta all distal of suggestion of articulation.
Antenna ( Figure 7 View FIGURE 7 E) six-articled, 0.6 times as long as antennule; article-1 short and annular, article-2 longer than broad, with dorsal seta; article-3 short, with dorsal seta; article-4 longer than articles 2–3 combined with four penicillate and two simple distal setae; article-5 short with long seta; article-6 very small, with four setae.
Labrum ( Figure 7 View FIGURE 7 F) subtriangular, as long as labium. Labium ( Figure 7 View FIGURE 7 G) two-lobed with slightly notched distal processes. Mandibles stout; right mandible ( Figure 7 View FIGURE 7 I) with finely crenulate distal margin and bifid incisor; molar weak and acutely pointed; left mandible ( Figure 7 View FIGURE 7 H) with crenulate incisor and broader lacinia mobilis, molar as in right mandible. Maxillule ( Figure 7 View FIGURE 7 J) endite sigmoid, with two distal setae and eight terminal spines. Maxilliped ( Figure 7 View FIGURE 7 K) basis with seta near articulation with palp; palp article-2 with three unequal weak setae, article-3 with three unequal setae and article-4 with four unequal simple setae and two stronger pinnate setae.
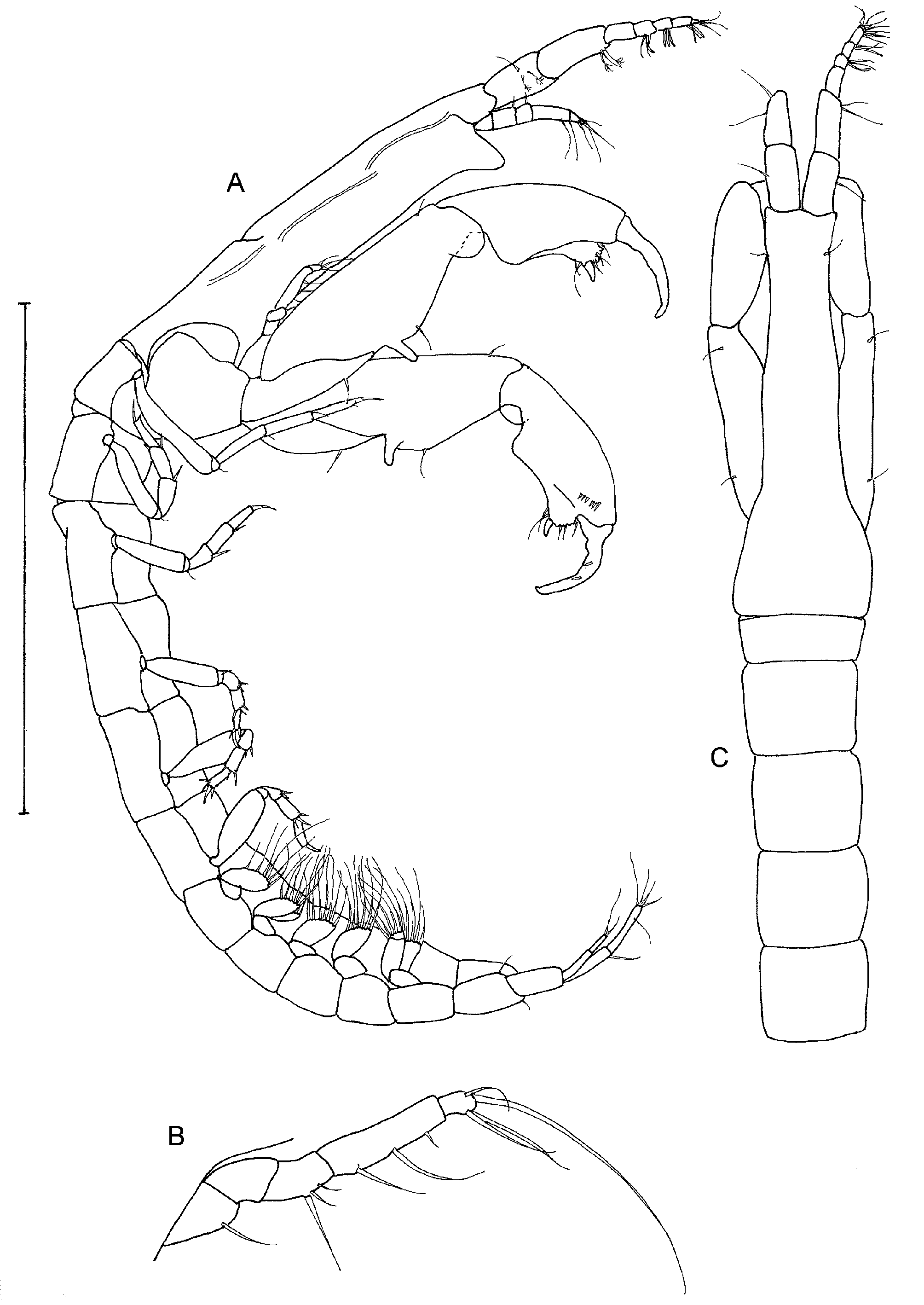
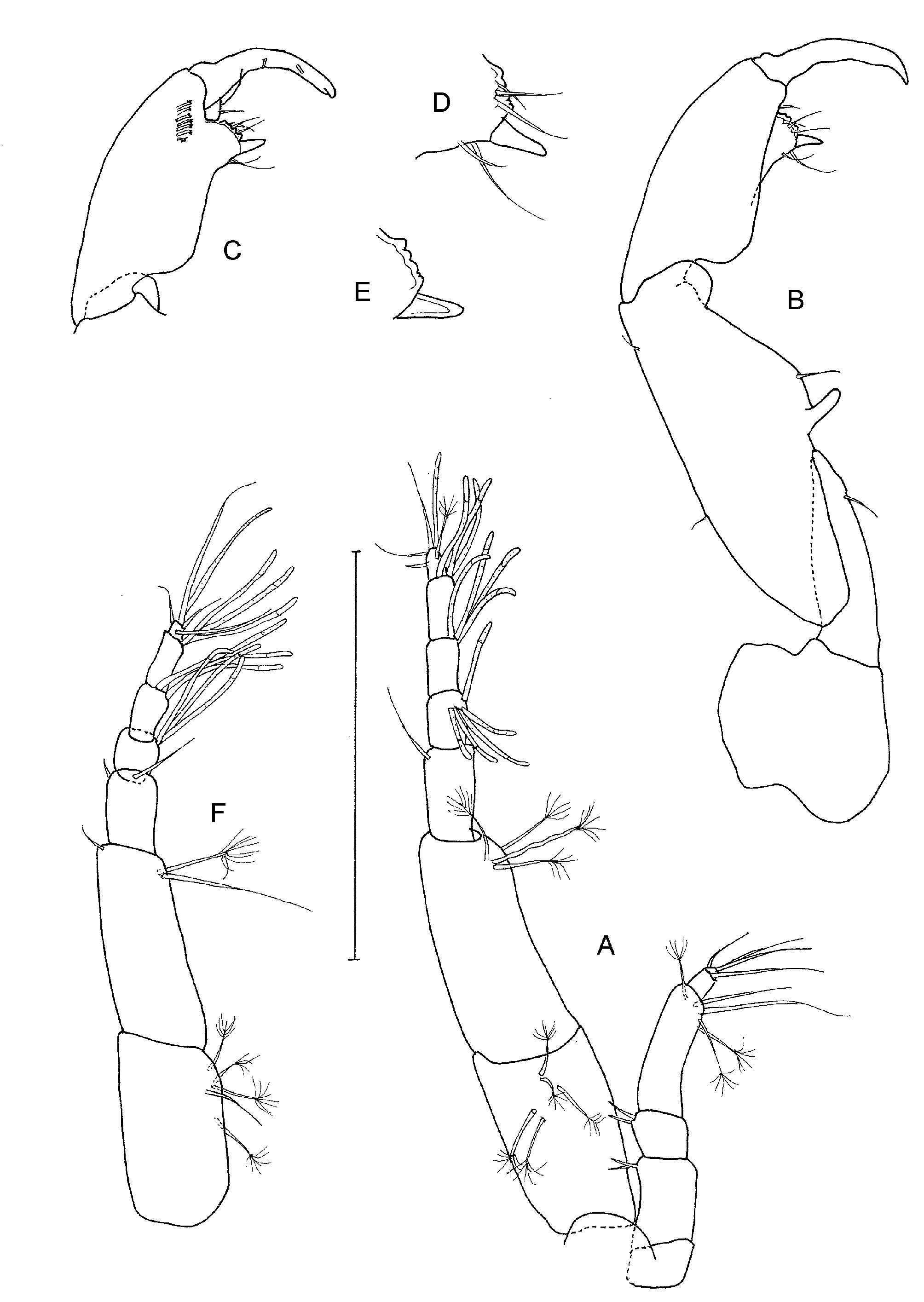
Cheliped ( Figs 7 View FIGURE 7 L, M) basis large, with rounded posterior free margin, attached to carapace only by a small dorso-posterior section, with small distolateral seta; merus subtriangular, with ventral seta; carpus 1.4 times as long as broad, with two dorsal and two ventral setae; chela 1.3 times as long as carpus, propodus with bifid dorsodistal crest and anterior comb of four dendritric setae ( Figure 8 View FIGURE 8 A) and associated small spines, fixed finger with weakly convex incisive margin and single distal subtriangular tooth, two unequal ventral setae, one seta near articulation with dactylus, and three unequal setae near incisive margin; dactylus narrow and acute, with several dorsal nodules and one small anterior seta.
Pereopod 1 ( Figure 8 View FIGURE 8 B) longer and more slender than pereopods 2–3; coxa with seta; basis five times as long as broad; ischium with small seta; merus nearly three times as long as broad, with small dorsal seta; carpus subrectangular, as long as merus, with three unequal setae, one as long as propodus; propodus 0.7 times as long as carpus, with small distal seta; dactylus and unguis 0.6 times as long as propodus, unguis spatulate at tip.
Pereopod 2 ( Figure 8 View FIGURE 8 C) relatively stout, coxa with seta; basis as long as merus, carpus and propodus combined, with dorsal penicillate seta; ischium with two small setae; merus distally expanded, 1.8 times as long as greatest width, with one seta and one spine; carpus about as long as merus, rectangular, 1.3 times as long as broad, with one dorsodistal spine, one larger ventrodistal spine and one very short spine, ventral margin with two spinules; propodus as long as carpus, but narrower, with distoventral spine and several ventral spinules; dactylus and unguis together as long as propodus, unguis spatulate at tip. Pereopod 3 ( Figure 8 View FIGURE 8 D) very similar to Pereopod 2 but basis slightly stouter, with dorsal simple seta.
Pereopod 4 ( Figure 8 View FIGURE 8 E) coxa indistinct; basis as in Pereopod 2 but without setae; ischium with two setae; merus 1.7 times as long as broad, expanded distally, with two ventral spines; carpus just shorter than merus, with four unequal spines and simple seta; propodus about as long as carpus, but narrower, with three distal spines and a few ventral spinules; dactylus and unguis fused, together longer than propodus. Pereopod 5 (not figured) similar to Pereopod 4. Pereopod 6 ( Figure 8 View FIGURE 8 F) similar to pereopods 4–5 but propodus with four distal spines and dactylus/unguis shorter than propodus.
Pleopods all similar ( Figure 8 View FIGURE 8 G), with basal article about as long as broad, naked; endopod and exopod rami similar, but exopod somewhat broader, with ten terminal setae, endopod with one distal and seven terminal setae; all setae plumose and longest barely as long as rami.
Uropod ( Figure 8 View FIGURE 8 H) slender, about twice as long as pleotelson; basal article three times as long as broad, as long as exopod; exopod two-segmented, as long as proximal article of two-segmented endopod; setation as figured.
Ovigerous female, generally similar to non-ovigerous female, but pereon dorso-ventrally compressed and with pairs of oostegites on pereopods 1 to 4; length 1.41–1.76 mm (cf. 2.18 mm T. lilljeborgi , 2.62–3.20 mm T. danica ).
Post-ovigerous females, generally similar to ovigerous female, but oostegites shed; length 1.38–1.79 mm.
Manca-II, generally similar to non-ovigerous female but pereonite-6 and pleonites shorter, lacking pereopod 6 and pleopods; length 0.68 mm. (cf. 0.99 mm T. lilljeborgi , 1.05 mm T. danica )
Manca-III, similar to manca-II but with rudimentary pereopod 6; length 1.00– 1.04 mm (cf. 1.3–1.4 mm T. lilljeborgi , 1.28 mm T. danica )
Juvenile, similar to manca-III but also with rudimentary (setae-less) pleopods; length 0.96–1.00 mm.
Terminal male, body ( Figure 9 View FIGURE 9 A) slender, length 2.83 mm (paratypes 1.87–2.3 mm). Cephalothorax ( Figure 9 View FIGURE 9 C) extremely attenuated, particularly rostral two-thirds that is almost conical, about three times as long as greatest width, as long as pereonites 1–4 and half of pereonite 5 combined; with dorsal ridges as in female and prominent conical process ventral to antennae; maxillipeds attached beneath caudal third of cephalothorax. Pereonites more or less rectangular, or with slightly convex lateral margins, pereonite-1 shortest, others about as long as broad, none elongate. Pleon about 1.5 times length of pereonites 5 and 6. Pleotelson short, only as long as preceding pleonite and basal article of uropods.
Antennule ( Figure 10 View FIGURE 10 A, F) seven-articled, half as long as cephalothorax; peduncle article 1 stout. 1.8 times as long as broad, with or without one simple lateral seta and at least four penicillate setae; peduncle article 2 longer than article 1, with one small medial seta, one long lateral seta and one to four penicillate setae; peduncle article 3 longer than any flagellar segments, with only two unequal setae; flagellum of four segments, proximal segment as long as broad, with group of five (or three in smaller paratype) aesthetascs; second segment slightly longer, with similar group of aesthetascs; third segment longer and more slender than second, with group of aesthetascs; distal segment small with at least four unequal simple and penicillate setae and one aesthetasc.
Antenna ( Figure 10 View FIGURE 10 A) just longer than first article of antennule peduncle, generally similar to that of female, but articles 2 and 3 with two dorsal setae.
Mouthparts absent apart from large maxillipeds. Maxilliped ( Figure 9 View FIGURE 9 B) basis with seta near articulation with palp; palp articles 1 and 2 similar in length, article 2 with two small and one long seta; article 3 longer than articles 1 and 2 combined, with two long setae and one small distal seta, article 4 small, with short and long terminal setae, as figured.
Cheliped ( Figure 10 View FIGURE 10 B) large and about as long as cephalothorax and antennules combined; attached at extreme caudal part of cephalothorax and posterior of basis overlaps sternal part of pereonite 1; basis with large free posterior margin but narrow articulation with merus; merus elongate, ‘conical’, with ventral seta; carpus slender, 2.5 times as long as greatest width, with finger-like ventral spur, one ventral and two dorsal setae, and small rounded distal shield on outer margin; chela almost as long as carpus; propodus ( Figure 10 View FIGURE 10 C, D, E) with prominent ventral process, one seta near articulation with dactylus and anterior comb of about seven strong spiniform setae; fixed finger twisted medially, short, with strongly convex incisive margin bearing five ‘teeth’, with three outer and two ventral setae, distal spine straight and conical; dactylus strongly curved, wide at base, with two inner peg-like spines.
Pereopod 1 ( Figure 11 View FIGURE 11 A) elongate, about 1.5 times as long as pereopods 2 and 3; coxa with seta; basis wider distally; ischium with seta; merus four times as long as broad, with ventral seta; carpus rectangular, as long and wide as merus, with stronger distal seta; propodus 0.75 times as long as carpus, with small distal seta; dactylus and unguis shorter than propodus.
Pereopod 2 ( Figure 11 View FIGURE 11 B) more slender than in female, basis, merus, carpus and propodus proportionately longer, setation similar but spines longer and basis with two penicillate setae. Pereopod 3 ( Figure 11 View FIGURE 11 C) similar to pereopod 2 but basis with single penicillate seta.
Pereopod 4 ( Figure 11 View FIGURE 11 D) generally similar to that of female but basis with penicillate seta, longer spines on merus, no simple seta on carpus and propodus proportionately longer. Pereopod 5 ( Figure 11 View FIGURE 11 E) similar to pereopod 4, but without basal penicillate seta. Pereopod 6 ( Figure 11 View FIGURE 11 F) similar to pereopods 4 and 5 but with four propodal spines.
Pleopods all similar ( Figure 11 View FIGURE 11 G), proportionately larger than in female and rami more elongate; endopod with one distal and seven terminal plumose setae, exopod with eight plumose setae, longest twice as long as rami.
Uropod ( Figure 11 View FIGURE 11 H) similar to that of female but basal article stouter.
Etymology: from Greek mikrothymos, ‘narrow-minded’, a pun on the male’s narrow cephalothorax and focus on detecting females for mating.
Distribution: throughout the survey area between 32º54.054’N 034º04.33’E and 31º41.328’N 034º33.326’E, in the shallow inshore waters ranging between 4.4 and 16.2 m depth.
Remarks. This small genus of slender nototanaids was partly revised by Bird (2002), who established that two distinct species were present in British and West European waters, and which are common in sandy substrata at intertidal and shallow-shelf depths (Bird op.cit., Holdich & Jones 1983). Differences in female cheliped and mandible molar structure, as well as male morphology (particularly pereon length, antennule and cheliped shapes), help distinguish the two previously described European species T. lilljeborgi and T. danica . Male antennule structure in T. lilljeborgi is of two long peduncular articles (owing to presumed fusion of the proximal two articles) and a four-segmented flagellum (terminal article small and indistinctly demarcated) all of which bear aesthetascs, whereas T. danica (males formerly known as T. elongatus Jones & Holdich, 1983 ) has a three-articled peduncle and four-articled flagellum with distinct small terminal article. The new species described above shares its antennule morphology with T. danica but has a pereon shape more similar to that of T. lilljeborgi . Elongation of the male cephalothorax, with associated reduction of mouthparts, is typical of several nototanaid genera such as Nototanais Richardson, 1906 and Nototanoides Sieg & Heard, 1985, but the extreme example shown by Tanaissus microthymus sp. nov. is comparable only with Paranesotanais longicephalus Larsen & Shimomura, 2008 .
At first glance, with their highly elongate cephalothorax and distinctive cheliped carpus structure, only the males of T. microthymus differ significantly from those of Tanaissus lilljeborgi , but the females are also separable, primarily on morphometrics, including the relative size of the pleon. In neuters and females the pleon is shorter than, or slightly longer than pereonites 5 and 6 combined (0.81–1.14 times as long), with a weak trend to lengthening with size, compared to ratios of 0.93–1.9 in T. lilljeborgi (with allometric increase) and 0.76–1.08 in T. danica (no allometric increase). The female cephalothorax is also slightly more elongate in T. microthymus than in T. lilljeborgi and the new species is also smaller than T. lilljeborgi and T. danica at equivalent life-cycle stages (see description above). The females of T. microthymus and T. lilljeborgi share cheliped morphology, with a bicuspid crest on the propodus, and this, together with more similar male morphology implies a closer phylogenetic relationship than with T. danica . Minor differences are, however, evident in pleopod shape, maxilliped palp, pereopod and pleopod setation; in addition, the anterior comb on the cheliped propodus is of dendritric setae in T. microthymus and the cheliped fixed finger of this species has only a single distal tooth.
It might be considered that the new species merely represents an East Mediterranean regional variant of the type species, but the differences, especially in male morphology, appear too large to accommodate this hypothesis. Remarkably, considering the huge number of specimens recorded off Israel in the present study (8633 specimens from 99 samples, densities up to 2,800 per m2), there are no previous records of the genus Tanaissus from the Mediterranean. This may well be a result of previous sample-analyses using too-coarse a mesh size. There is a typically enigmatic record by Makkaveeva (1971) of “ Tanaissus lilljeborgi ” from the Red Sea, which may in fact be the present species. Ideally, Tanaissus populations from regions westwards around the Mediterranean, through to the Iberian Atlantic seaboard should be studied, along with DNA molecular analyses, to establish true phylogenetic affinities and relationships.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.