
Ceraticelus phylax Ivie & Barrows, 1935
|
publication ID |
https://doi.org/ 10.5281/zenodo.190335 |
|
DOI |
https://doi.org/10.5281/zenodo.6225643 |
|
persistent identifier |
https://treatment.plazi.org/id/997CC11F-FFCB-010B-FF78-196B3F22E2EE |
|
treatment provided by |
Plazi |
|
scientific name |
Ceraticelus phylax Ivie & Barrows, 1935 |
| status |
|
Ceraticelus phylax Ivie & Barrows, 1935 View in CoL
( Figures 1–13 View FIGURES 1 – 6 View FIGURES 7 – 10 View FIGURES 11 – 16 , 29 View FIGURE 29 )
Ceraticelus phylax Ivie & Barrows, 1935: 13 View in CoL , pl. 5, figs. 33–36.
Types. Ceraticelus phylax Ivie & Barrows View in CoL , holotype male and allotype female presumed lost or misplaced. Type locality: U.S.A.: Oklahoma, Chickasha. Male holotype, female allotype; coll. M. W. Shackleford, June 8, 1934. Originally deposited in the University of Utah collection.
Material examined. U.S.A.: California: Riverside County: Riverside (coordinates indicate San Diego Co., NE of Poway), [33°N 117°W] 23, April–May 1962, coll. E. I. Schlinger (det.; V. Roth, ver.: W. Ivie, 1962) (coll. AMNH); Santa Rosa Plateau, Sylvan Meadows, 570 m [33°32'N 117°17'W] 2Ƥ, 7 July 1999, sweepnet, coll. C. Dunning; Santa Margarita Ecological Preserve, near north entrance, 349.3 m [33°27'43"N 117°10'11"W] 2Ƥ, 2 June 1997, vacuum sample California sagebrush ( Artemisia californica ), coll. R. Redak lab & T. Prentice; San Diego County: Miramar Naval Air Station (now, Marine Corps Air Station Miramar): NE corner of base near W Sycamore Canyon, 290 m [32°55'03.0"N 117°00'10.6"W] 10Ƥ, 20 August 1998, 33 7Ƥ, 22 June 1998 ( CAS: 13 (9032940) 1Ƥ (9032941); AMNH: 13 1Ƥ); near NE corner of base between W Sycamore Canyon & Sycamore Canyon, 300 m [32°55'16.6"N 117°59'43.2"W] 1Ƥ, 22 June 1998, vacuum samples coastal sage scrub, coll. R. Redak lab & T. Prentice; Marine Corps Base Camp Pendleton: Alfa Two, coastal side of base, 47 m [33°22'49.3"N 117°33'01.0"W] 13 3Ƥ, 17 May 1996, 13 1Ƥ, 26 August 1995; SW corner of base N of San Onofree Creek & Basilone Rd., 37.2 m [33°23'23.8"N 117°33'17.2"W] 23 2Ƥ, 13 May 1996, 13 1Ƥ, 21 August 1995, 13, 16 May 1995; Romeo Three, coastal side of base, 81.7 m [33°21'22.1"N 117°31'33.7"W] 13 1Ƥ, 13 May 1996; Juliette, east side of base, 138.7 m [33°19'31.7"N 117°17'40.3"W] 1Ƥ, 10 May 1996; Alfa One, 133.8 m [33°24'53.9"N 117°31'49.3"W] 1Ƥ, 26 August 1995, 13, June 1994; Uniform, coastal side of base near beach, 36.3 m [33°19'18.9"N 117°29'11.2"W] 1Ƥ, 26 August 1995; Romeo Two, coastal side of base, 81.4 m [33°20'58.6"N 117°30'52.4"W] 1Ƥ, 19 August 1995; Papa Three, S of Basilone Rd., 214.6 m [33°22'18.6"N 117°27'16.0"W] 1Ƥ, 14 August 1995, 3Ƥ, 23 May 1995, 13, June 1994; Romeo Two, coastal side of base, 69.2 m [33°21'02.0"N 117°31'03.6"W] 1Ƥ, 14 August 1995; Hotel, east side of base, 206.4 m [33°24'13.1"N 117°17'50.6"W] 1Ƥ, 8 June 1995; Romeo Three, coastal side of base, 55.2 m [33°21'50.8"N 117°32'22.4"W] 1Ƥ, 30 May 1995; Romeo One, S of Basilone Rd., 246.6 m [33°22'36.6"N 117°26'17.3"W] 13, 23 May 1995; Alfa Two, coastal side of base, 92.7 m [33°22'03.7"N 117°32'26.8"W] 1Ƥ, June 1994, vacuum samples coastal sage scrub, coll. R. Redak lab & T. Prentice. Texas: Hidalgo County: 11.3 km (7 mi) NW of Edinburg, [98°W 26°N] 2Ƥ (only one with epigynum), 29 December 1936, coll. Stanley Mulaik (det.: W. Ivie) (coll. AMNH).
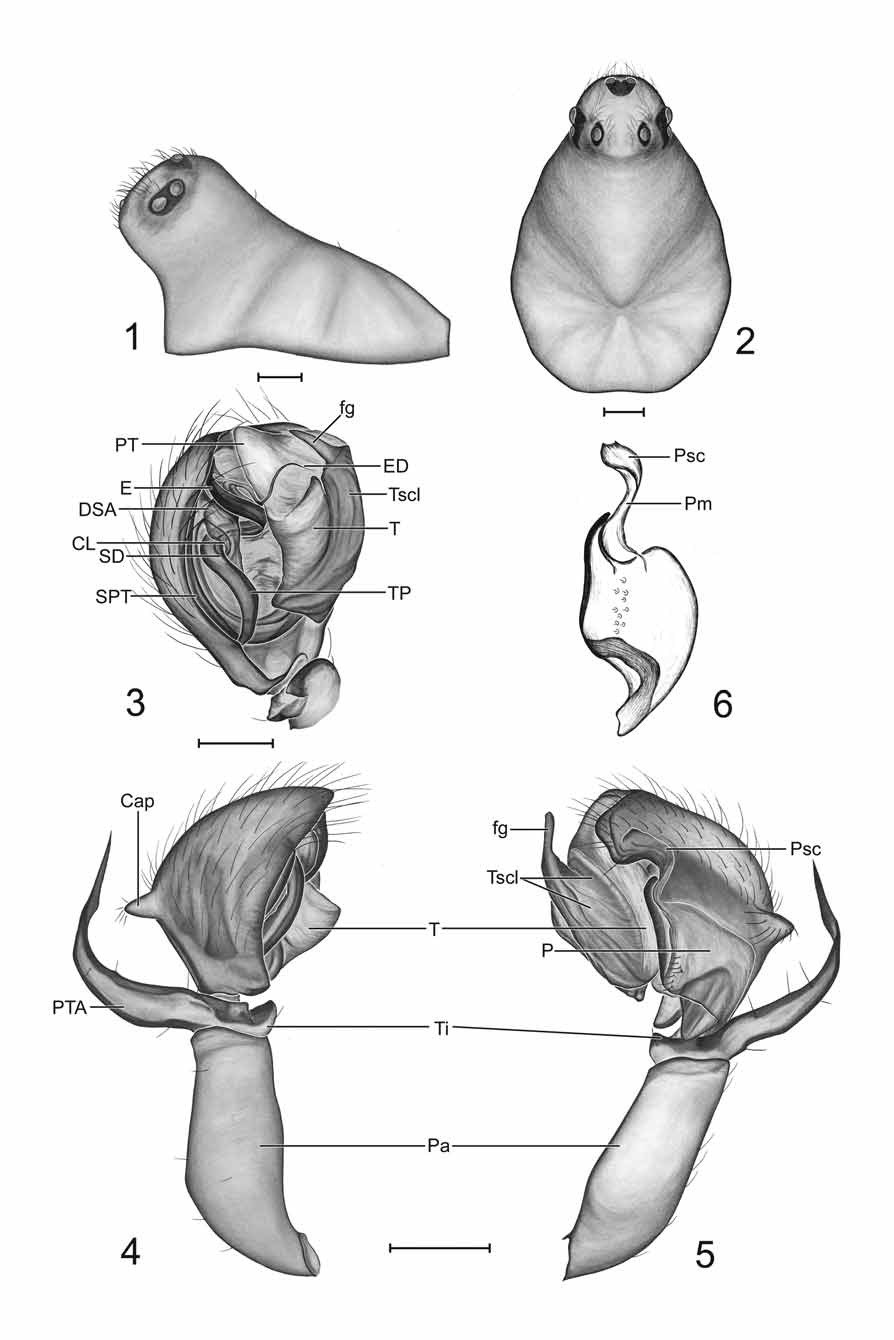
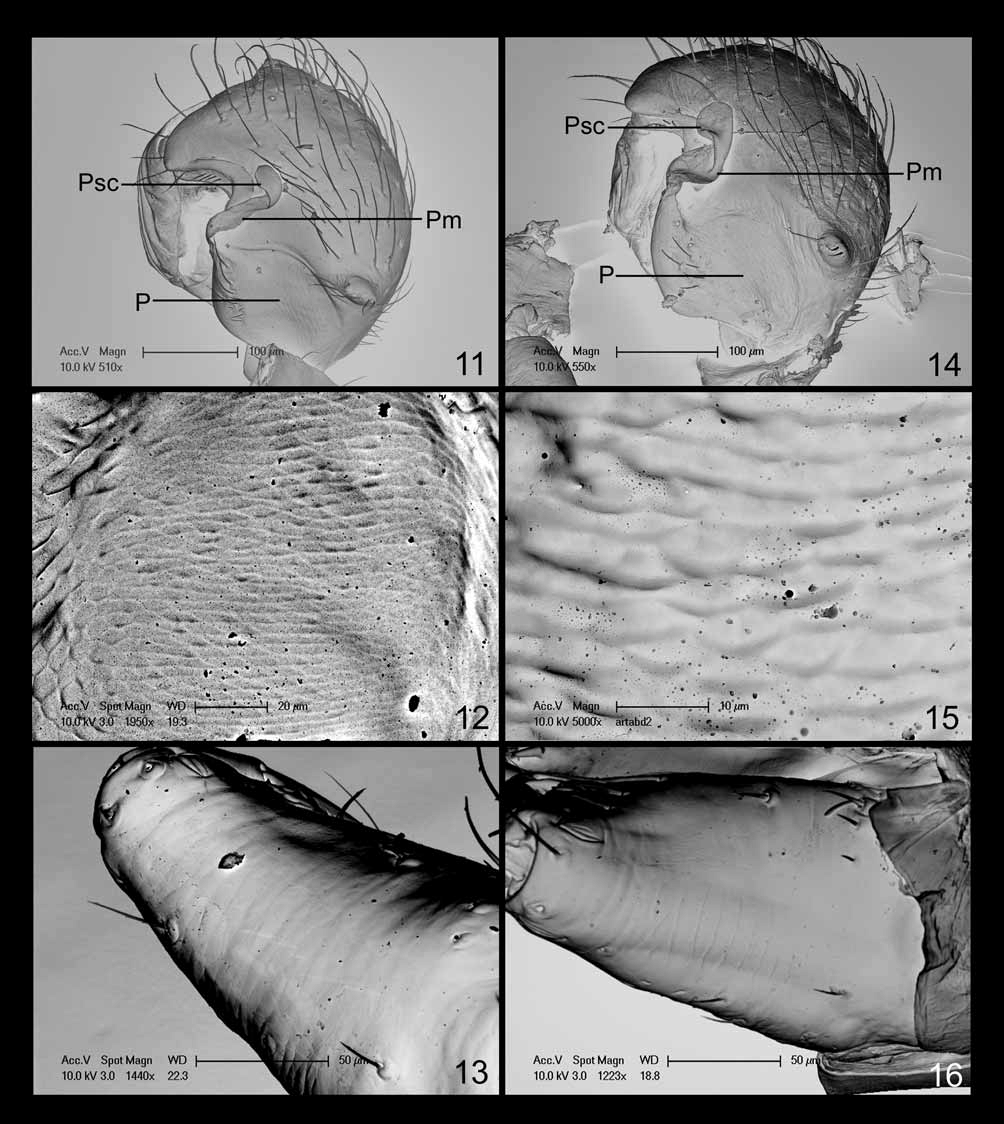
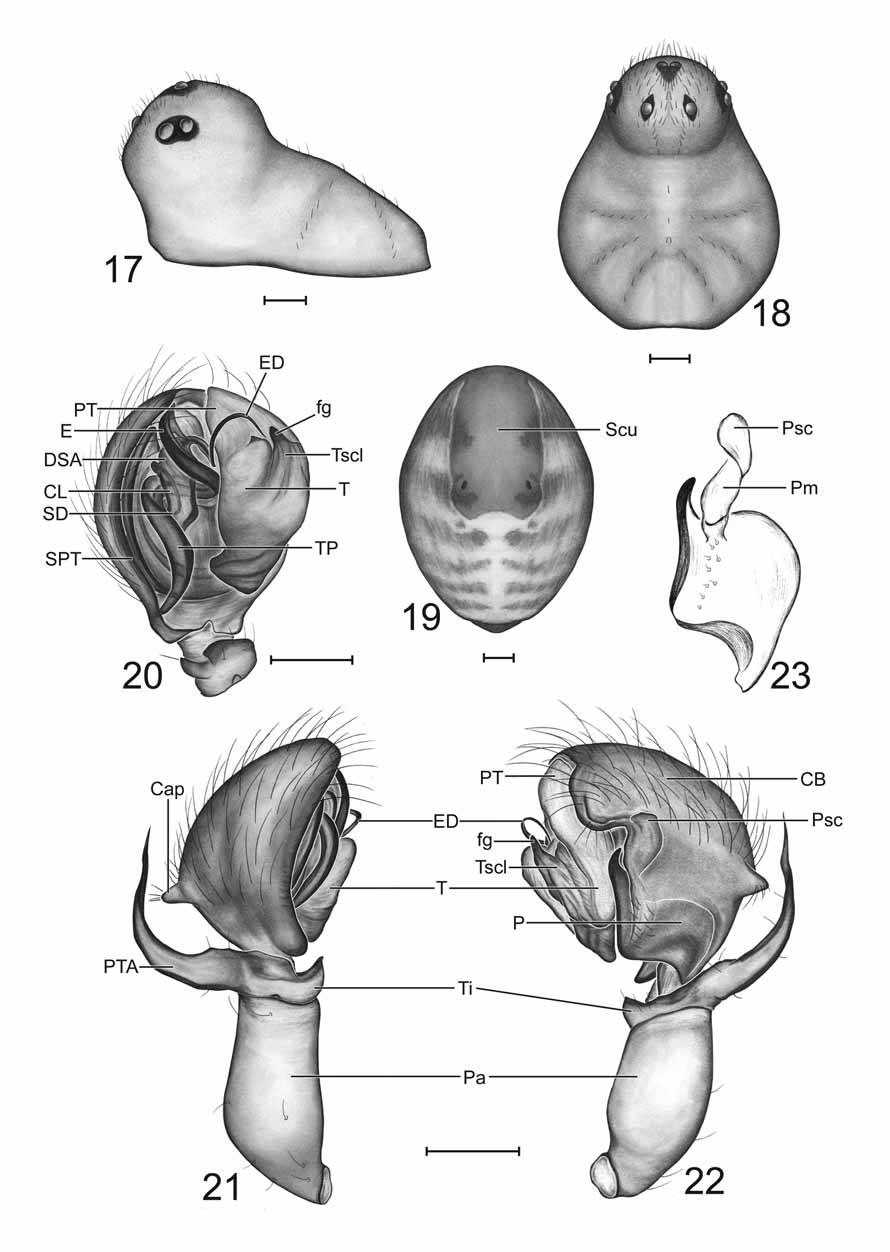
Diagnosis. Males of Ceraticelus phylax can be distinguished from those of all other species except C. limnologicus Crosby & Bishop by the height and narrowness of the steeply projecting cephalic lobe ( Figs 1, 2 View FIGURES 1 – 6 ) and from males of C. limnologicus by the small angular protrusion of the cymbium (Cap: Fig 4 View FIGURES 1 – 6 ) which is large and rounded in C. limnologicus (Fig 62: Crosby & Bishop 1925) and the wide sigmoid shape of the paracymbium (P: Figs 5, 6 View FIGURES 1 – 6 , 11 View FIGURES 11 – 16 ) which is long, slender, and tapering in C. limnologicus ( Crosby & Bishop 1925: 32) . Males can be further distinguished from those of C. artemisiae sp. nov. by the longer distal finger of the tegular sclerite (fg: Figs 3, 5 View FIGURES 1 – 6 ; compare to Figs 20, 22 View FIGURES 17 – 23 ) and absence of abdominal markings.
Females can be distinguished from those lacking a dorsal scutum with similar epigynal morphology, C. atriceps (Cambridge) , C. silis Dondale , C. similis (Banks) , and C. artemisiae sp. nov., by the relative position and form of the spermathecae and copulatory tubes in ventral view ( Fig 8 View FIGURES 7 – 10 ; compare to Figs 799, 832, 839: Paquin & Dupérré 2003 and Fig 26 View FIGURES 24 – 28 , respectively), and also from C. atriceps and C. artemisiae sp. nov. in dorsal view by the convolution of the copulatory tubes ( Fig 10 View FIGURES 7 – 10 ; compare to Fig 6 View FIGURES 1 – 6 : Levi & Levi 1955) in the former species and by the smaller loops of the copulatory ducts and the position of the anterior bends of the ducts relative to the spermathecal apex ( Fig 10 View FIGURES 7 – 10 ; compare to Fig 28 View FIGURES 24 – 28 ) in the latter species. Females are further distinguished from those of C. artemisiae sp. nov. by the absence of abdominal markings and the entirely darkened eye region ( Fig 7 View FIGURES 7 – 10 ; compare to Fig 24 View FIGURES 24 – 28 ).
Description. Males (n=8). Total length ~1.55–1.80. Carapace: length 0.81–0.86 (mean 0.83), width 0.59– 0.63 (mean 0.62), light brownish to yellowish orange with narrow marginal black or dusky border, pars thoracica with fine reticulate microsculpture, without cervical constriction, outline continuing around lower clypeal region in circular fashion, narrower in front, somewhat egg shaped, margin interrupted in dorsal view by cephalic projection, rising flatly or slightly convexly to level of coxa I, then rising concavely to apex of lobe. Cephalic region: upper portion of cephalic lobe occasionally lighter in color than remainder of carapace, lobe steeply projecting anteriad lower anterior margin of carapace, high and narrow in profile ( Fig 1 View FIGURES 1 – 6 ; also see Fig 33: Ivie & Barrows 1935), convexly descending from apex to flattened area just anterior to PME then descending concavely below most anterior convex point to lower clypeal margin; setae on each side between AME and PME directed medially, setae below AME usually directed anterodorsally; cephalic height 0.48– 0.52, cephalic width 0.28–0.31, cephalic width/carapace width 0.46–0.49 (mean 0.48), clypeal height 0.31– 0.38. Eyes: ocular area situated around top of cephalic lobe, PME ~2.0–2.8 diameters apart, either slightly anteriad or comprising highest point of lobe, PME–PLE ~0.65–1.0X PME diameter, AME relatively close together (~0.5–1.0 diameters apart) on flattened slope of anterior descending portion of lobe, ocular region usually darkened (sometimes lightly so) with black rings surrounding eyes, PME usually slightly larger than or occasionally subequal to contiguous and subequal PLE and ALE (ALE often slightly larger), AME smallest, AER slightly to moderately recurved, PER straight to slightly recurved. Chelicerae: not robust, lateral surface usually slightly concave, stridulatory striae very weakly developed ( Fig 13 View FIGURES 11 – 16 ) (probably nonfunctional), palpal plectra not detected (striae detectable on cleared chelicerae at 400X magnification under transmitted light), fangs well developed, promarginal teeth 4–6, 5 most common, retromarginal denticles 2–4. Sternum and pedicel: sternum yellowish orange, wider than long, widest between coxae I & II with margin slightly projecting between coxae, widely truncate between coxae IV, posterior width subequal to length of coxa IV, dusky around margin; pedicel sclerotized, ventral sclerite surrounding ventral, lateral, and dorsolateral surfaces, dorsal sclerite narrow, separated from ventral sclerite on each side by narrow membranous lines. Abdomen: immaculate, with yellowish orange setiferous dorsal scutum, varying greatly in degree of development (both in width and length), shorter and narrower than length and width, respectively, usually extending over anterior edge but distinctly short of reaching epigastic sclerite, setal bases in soft regions surrounded by diminutive sclerites, soft areas pale, yellowish-white, all ventral sclerites yellowish orange, epigastric sclerite heavily sclerotized, widely surrounding pedicel, laterally extending slightly distad of epigastric furrow, posterolateral edges curving mesally, epigastric plates with imbricated stridulatory striae ( Fig 12 View FIGURES 11 – 16 ) (visible after dissection and clearing using compound microscope at magnification of 200X), cuticular stridulatory plectra on posterodistal corners of hind coxae; inframammillary sclerite more weakly sclerotized than epigastric sclerite, confined to ventral surface. Legs: lighter than carapace, more yellowish to yellowish-white, leg I longer than leg IV, length femur I (LFI) usually slightly greater than carapace width (CW), occasionally (in one male) equal, length leg I (LLI) to carapace width (LLI)/CW) 3.40–3.65, tibia I longer than metatarsus I, Tibia I (TiI) l/d 6.8–7.5, tibial spines absent, TmI 0.51–0.56, TmIV absent. Palp: patella long, more robust than femur but slightly shorter, dorsolaterally swollen in proximal third ( Fig 5 View FIGURES 1 – 6 ) resulting in a ventromedial orientation of patella, length patella 0.35–0.39, length femur 0.41–0.43, tibia short, tibial apophysis extending laterally (horizontal portion), bending apically at right angle just slightly past midpoint, more or less parallel sided until beginning of bend then attenuate (vertical portion) through bend to sharp terminus (PTA: Fig 4 View FIGURES 1 – 6 ); tibial trichobothria not detected. Palpal bulb: cymbium with small dorsolateralsubbasal angular protrusion (Cap: Figs 4, 5 View FIGURES 1 – 6 ), broadest mesolaterally with single transverse row of laterally curving setae ( Figs 4, 5 View FIGURES 1 – 6 ); paracymbium forming proximal half of ventrolateral outline of cymbium, with incomplete fission from cymbium (refer to introductory comments in this section), more or less widely sigmoid in shape (left palp viewed with anterior end up) ( Figs 5, 6 View FIGURES 1 – 6 , 11 View FIGURES 11 – 16 ) with narrow apical free darkened finger, greatly widening through curves and narrowing proximally; membrane extending from near base of finger, terminating in scale-like structure adhering to the cymbium (Pm, Psc, respectively: Figs 6 View FIGURES 1 – 6 , 11 View FIGURES 11 – 16 ); suprategulum with a short sclerotized distal suprategular apophysis (DSA: Fig 3 View FIGURES 1 – 6 ), notched on its ventroectal margin, with base of ejaculatory duct apparently lying within notch and supported by the structure; column sclerotized, somewhat more lightly so than suprategular apophysis (CL: Fig 3 View FIGURES 1 – 6 ); radical division of bulb with long spiraled tailpiece curving into and terminating mesobasally in alveolus (TP: Fig 3 View FIGURES 1 – 6 ); embolic division spiraling apically ~one and a quarter turns with embolic ribbon terminating dorsally at distal edge of mesal side of cymbium, ejaculatory duct spiraling off embolic ribbon preapically in ventral direction for ~one and a half turns, apex resting in cradle between tegulum and protegulum just behind base of finger of tegular sclerite (ED: Fig 3 View FIGURES 1 – 6 ; also see Fig 35: Ivie & Barrows 1935); tegular sclerite (Tscl) with distal finger elongated (fg: Figs 3, 5 View FIGURES 1 – 6 ; also see Figs 34, 35: Ivie & Barrows 1935).
Females (n=12). Total length ~1.5–1.85. Carapace: length 0.63–0.74 (mean 0.70), width 0.52–0.59 (mean 0.57), color and dusky marginal band as in male, with very slight cervical constriction at most, in profile rising most abruptly posteriorly at relatively flat incline to level between coxae II & III then gradually to highest point just behind or at level of PME ( Fig 7 View FIGURES 7 – 10 ), pars thoracica with fine reticulate micosculpture, cephalic height 0.27–0.31, ocular width/carapace width 0.44–0.50 (mean 0.46), clypeal height 0.13–0.16. Eyes: AER & PER usually straight sometimes slightly recurved, PME ~0.7–1.0 diameters apart, PME–PLE ~0.45–0.60X PME diameter, PME and ALE usually subequal in size, PLE usually slightly smaller, PEs contiguous, AME smallest, ~0.45–0.70 diameters apart, black rings around eyes, eye region darkened ( Fig 7 View FIGURES 7 – 10 ). Chelicerae: more stout than in male, stridulatory striae not detected (on cleared chelicerae at 400X magnification under transmitted light), 4–5 promarginal teeth, 5 most common, 2–4 retromarginal denticles, 3 most common. Sternum and pedicel: sternum usually as in male although length occasionally equals width, pedicel as in males. Abdomen: immaculate, lacking dorsal shield, muscle impressions with rounded sclerites distinctly larger than diminutive sclerites surrounding setal bases, soft areas pale, yellowish-white, coloration of ventral sclerites as in males, epigastric sclerite well developed widely surrounding pedicel, membranous lines separating median portion from lateral, epigastric plate stridulatory striae, coxal plectra, sclerites of pedicel, and inframammillary sclerite as in males. Legs and palps: coloration of legs as in male, length leg I less than, equal to, or greater than length leg IV, LLI/CW 3.24–3.62, LFI/CW 0.92–1.06, tibia I longer than metatarsus I, TiI l/d 5.7–6.7, tibial spines absent, TmI 0.45–0.59, TmIV absent, palpal tibia with 2 trichbothria. Epigynum: with laterally positioned spermathcae usually curved toward each other like parentheses, directed anteriad to slightly anteromediad (S: Fig 8 View FIGURES 7 – 10 ); dorsal plate triangular in shape (DP: Fig 9 View FIGURES 7 – 10 ), narrowing anteriorly, anterior edges forming medial borders of genital openings, in ventral view plate often appearing as an inverted ‘T’ with posterior transverse portion appearing either rectangular or triangular (DP: Fig 8 View FIGURES 7 – 10 ); copulatory openings beneath internal darkened semicircular ridges that extend from beneath anterior portion of dorsal plate to the posterolateral edges of the plate near base of spermathecae ( Fig 8 View FIGURES 7 – 10 ; also see Fig 36: Ivie & Barrows 1935); copulatory ducts leaving openings in dorsolateral direction, travelling anteriorly then turning toward the ventral surface slightly posteriad of or at level of apex of spermatheca in a tight posteromesally directed loop (CD: Fig 10 View FIGURES 7 – 10 ), continuing posteriorly then anteriorly in a loose U-shaped loop (CD: Figs 8, 10 View FIGURES 7 – 10 ; also see Fig 36: Ivie & Barrows 1935), forming the second tight loop (usually at the same level as the first, sometimes slightly posteriad) that curves dorsally and then proceeds posteriorly, finally emptying into base of spermatheca dorsomesally (CD: Fig 10 View FIGURES 7 – 10 ).
Distribution. Specimens are known from Riverside and San Diego counties in southern California ( Fig 29 View FIGURE 29 ), Chickasha, Oklahoma, and from near both Edinburg, Texas and Lincoln, Nebraska. The species’ known California distribution is closely aligned with that of C. artemisiae sp. nov. (see Prentice et al. 1998: 188, 195). Thus far, in all regions from which we have collected specimens, C. artemisiae sp. nov. also occurs. The earliest known specimens from California were collected in 1962 from Riverside County (although coordinates on the collection label indicate San Diego County near Poway).
If the establishment of C. phylax in California is considered a range extension, it is significant given that the nearest known collection locality (Chickasha) is approximately 1830 km to the east. Lincoln, Nebraska and Chickasha, Oklahoma are separated by only 600 km and much of the area around Chickasha is very similar ( Oklahoma Atlas Institute 2005) to the prairie landscape description provided by Muma & Muma (1949) for the collection locality near Lincoln. Edinburg, Texas is approximately 1030 km south of Chickasha but most of the area surrounding Edinburg is classified as cropland ( Walter Geological Library 2000). Theridion llano Levi (Theridiidae) and the Argiope blanda O. P.-Cambridge ( Araneidae ), specimens of which were collected during our 1994–1996 study ( Prentice et al. 1998), were new California species records; the nearest collection localities of both species are within 75 km of Edinburg, Texas, approximately 1960 km to the east of San Diego. Since regular and/or reserve military bases are found in relatively close proximity to collection sites of C. phylax in Nebraska and Oklahoma and to sites in Texas where all three species are found, it seems more plausible that their occurrence in California resulted from incidental introduction (and possibly of C. phylax into southern Texas) via military personnel movement than from natural dispersal events over such significant distances. Additionally, because it seems highly unlikely that two sympatric probable sibling spider species would naturally co-occur, especially in pristine vegetation communities, in the absence of complete habitat partitioning, we suggest that C. phylax is an advent to southern California and that the species is more likely endemic to the prairieland between Nebraska and Texas.
Habitat. In California Ceraticelus phylax is primarily known from coastal sage scrub communities (see Prentice et al. 1998: 188, 195 – 96). Sixty percent (18) of the specimens collected in vacuum samples at MCBCP were from plots dominated or co-dominated by Artemisia californica and ninety-five percent (20) of those collected at MCASM were from a single plot dominated by California Broom ( Lotus ) and dense annual grasses. Two females were taken in vacuum samples from California sagebrush ( Artemisia californica ) in southwestern Riverside County (also refer back to ‘Introduction’). Specimens from near Lincoln, Nebraska were collected from virgin prairieland dominated by big bluestem, blue grama, little bluestem, and Kentucky bluegrass ( Muma & Muma 1949). The habitats of the type specimens from Chickasha, Oklahoma and the females collected in Edinburg, Texas are not specifically known (refer back to ‘Distribution’ section).
Phenology. Males have been collected in California from April to August and females from May to August (see Prentice et al. 1998: 188). One penultimate male was also taken in August samples, an indication that mature specimens are likely found in California somewhat later in the year. The two females from Edinburg, Texas were collected in late December. Adults from prairieland near Lincoln, Nebraska occurred only in July and August samples ( Muma & Muma 1949).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ceraticelus phylax Ivie & Barrows, 1935
| Prentice, Thomas R. & Redak, Richard A. 2009 |
Ceraticelus phylax
| Ivie 1935: 13 |