
Meridionale harrisi ( Arango and Brenneis, 2013 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3765.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:81FEEBE5-46BE-4AFC-A731-7D01D298E87B |
|
DOI |
https://doi.org/10.5281/zenodo.6128045 |
|
persistent identifier |
https://treatment.plazi.org/id/A033520D-FFB5-FD63-FF35-975E8CC8B693 |
|
treatment provided by |
Plazi |
|
scientific name |
Meridionale harrisi ( Arango and Brenneis, 2013 ) |
| status |
|
Meridionale harrisi ( Arango and Brenneis, 2013) View in CoL
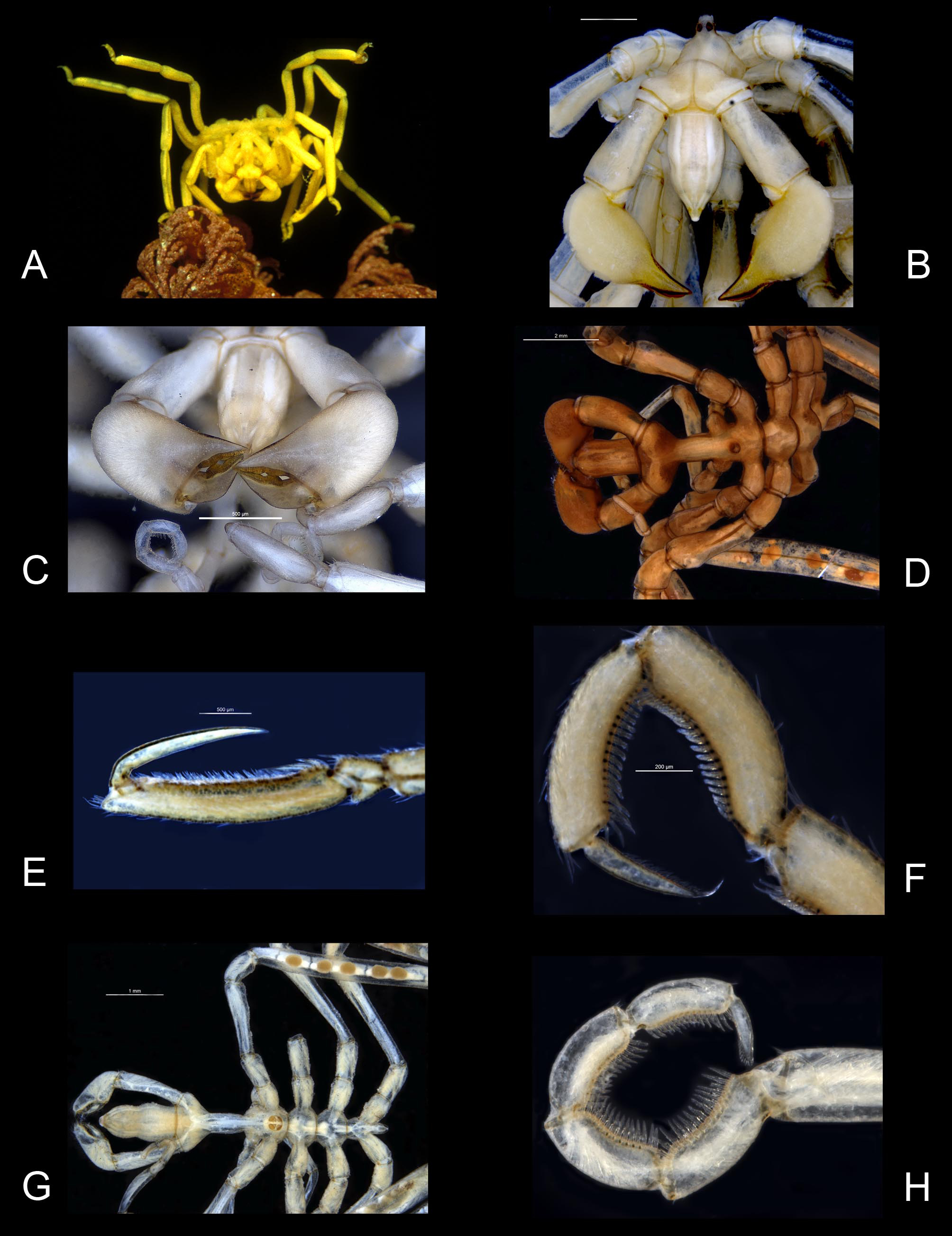
Figures 4 View FIGURE 4 B, C
Pseudopallene harrisi Arango and Brenneis, 2013 View in CoL
Remarks. Because of its distinctive markings, M. harrisi is somewhat of an iconic Australian east coast species, being much photographed since the early 1970s. The most northerly record of this colour form known by the author is Heron I., Queensland, where it was recorded swimming, and the most southerly record is Montague I. off the New South Wales southern coast which is very close to the type localities of M. laevis and M. ambigua . Contrary to the perception that pycnogonids have a limited dispersal capacity, long-legged pycnogonids such as those belonging to the Callipallenidae , Nymphonidae , Colossendeidae and Phoxichilidiidae are all efficient ‘swimmers’ and capable of drifting in the water column, potentially over wide distances. The migration of eggbearing males thereby ensures ongoing gene flow along the Australian coastline. The absence of records of M. harrisi south of Montague I. is therefore puzzling. The possibility of this being a consequence of a phylogeographic break in the south-east region is not supported by the geographic ranges of other species, often in different families, that significantly overlap this region. Meridionale reflexa for example is recorded from southern Tasmania , South Australia and Bass Strait to Moreton Bay in Queensland. My suspicion is that M. harrisi is present in Bass Strait in another colour form, probably predominantly yellow and possibly with red markings.
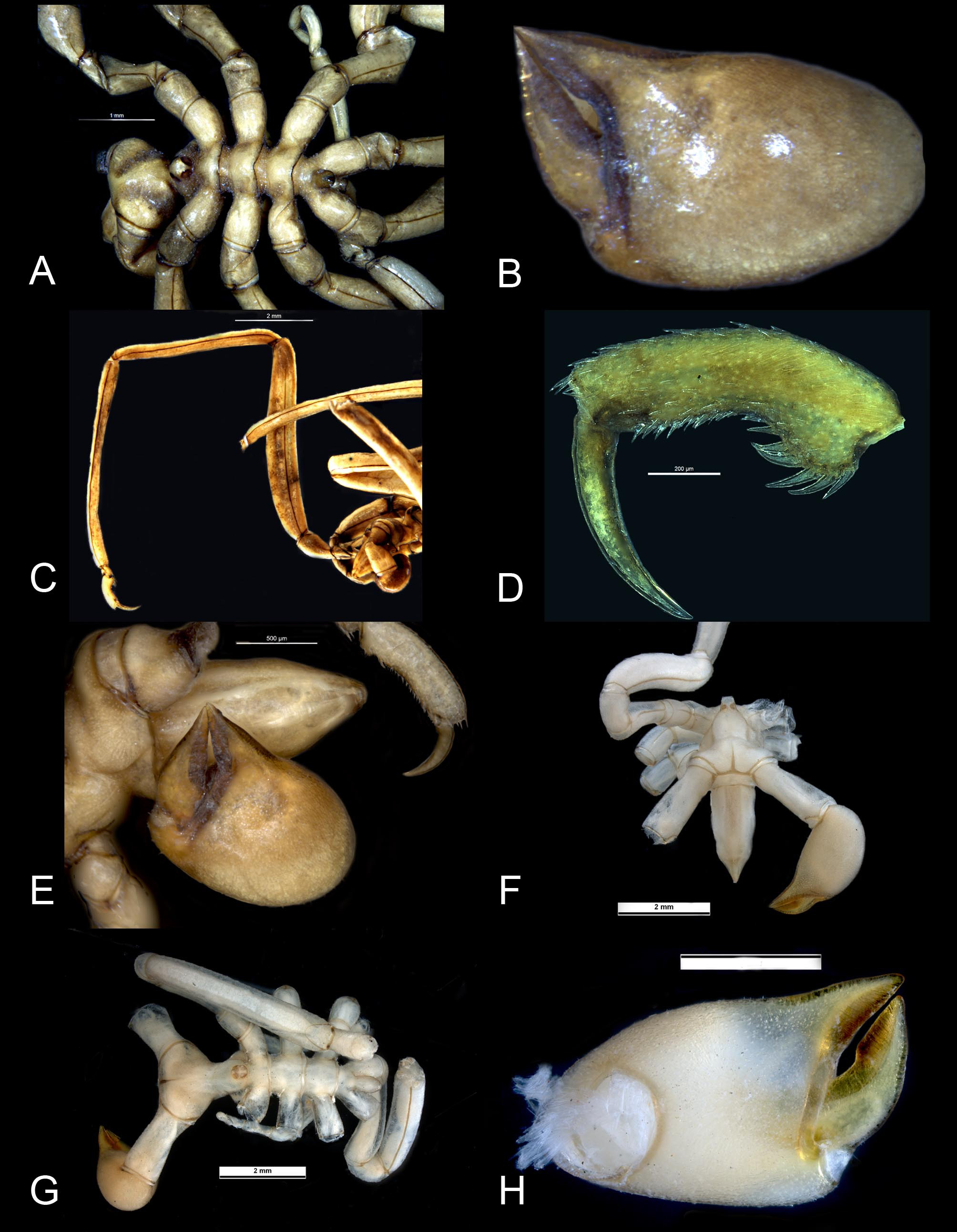
Based on the summary of distinguishing morphological features ( Arango & Brenneis 2013, Table 3) there is little to separate M. harrisi from M. ambigua . Unfortunately the distinguishing characteristics of each species were addressed inconsistently making it difficult to establish points of difference. The observation of a variable neck length in M. harrisi is particularly confusing. The prominence of the pre-ocular dorsal mound figured by Arango & Brenneis (fig 13C), also appears to be variable. Based on specimens of M. harrisi from Port Macquarie and Montague I. and specimens of P. ambigua from Bass Strait, there is no discernable difference in the mound shapes ( Figs. 3 View FIGURE 3 F, 4B). Arango and Brenneis distinguish Meridionale harrisi by its larger body size, a slightly more robust appearance, the number of oviger spines and the denticulations on the oviger claw. Based on Stock’s (1956a) measurements, the body length of the M. ambigua holotype (5.6 mm) is in fact longer than that of M. harrisi (5.35) and it is now established that the remaining oviger claw of the M. ambigua holotype is worn or damaged and cannot be compared to other species. Intraspecific variation in oviger and heel spine numbers is also not unusual and is indicative of pycnogonids generally. The oviger spine formula of the M. harrisi holotype is 16:13:11:11 compared to 14:12:11: 10 in the specimen from Port Macquarie and although the holotype is described as having four heel spines, three spines appear to be common. Despite the apparent differences in the proboscis shapes of the M. ambigua and M. harrisi holotypes ( Figs 3 View FIGURE 3 F, 4B) no differences were evident in additional specimens of both species. A possible explanation is that the proboscis of the preserved M. ambigua holotype is extended, exposing the broad arthrodial membrane ( Fig 3 View FIGURE 3 F). When retracted, the proboscis would not only appear to be shorter but the first constriction around the proboscis would almost be concealed by the anterior margin of the cephalon. Should this be the case, there would be little to distinguish the proboscides of the holotype specimens. The shapes of the immoveable fingers show some variability even within the same specimen. In the Montague I. specimen the left immoveable finger is evenly curved throughout its length as in the M. ambigua holotype, whereas the right immoveable finger has a stronger bend (or elbow) a little before its mid-length, more in keeping with the M. harrisi holotype, although this too is variable ( Fig 4 View FIGURE 4 C). Both chela finger shapes are found amongst Bass Strait material but variability in the cuticular cutting margins make it difficult to assign the specimens to one species or the other. The typical M. ambigua chelae form ( Fig. 3 View FIGURE 3 H) is very common in Bass Strait but due to the lack of sufficient material it is not possible to make the same assessment of M. harrisi specimens in waters north of Montague Island.
For the time being I have retained the independence of both species based on the distinctive colour pattern of M. harrisi , particularly given the close geographic proximity of the two colour forms, the as yet unresolved variability of the chelae shapes found in Bass Strait and the need to sight a wider range of specimens from north of Montague Island.
Distribution Montague I. to Heron I. Queensland.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Meridionale harrisi ( Arango and Brenneis, 2013 )
| Staples, David A. 2014 |
Pseudopallene harrisi
| Arango and Brenneis 2013 |