
Elysia pratensis Ortea & Espinosa, 1996
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:91353147-FDA8-45CC-A8F1-1DE801C835A6 |
|
DOI |
https://doi.org/10.5281/zenodo.5664211 |
|
persistent identifier |
https://treatment.plazi.org/id/A04A7E6D-9C1E-FFD6-46C9-FA00FEDD1A93 |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia pratensis Ortea & Espinosa, 1996 |
| status |
|
Elysia pratensis Ortea & Espinosa, 1996 View in CoL
( Figs. 6 View FIGURE 6 R, 48–50)
Elysia pratensis Ortea & Espinosa 1996: 116 View in CoL –119, figs. 1–3 — Redfern 2001: 163, figs. 675; Espinosa et al. 2005: 56; Valdés et al. 2006: 70 –71; Krug 2009: table 2, figs. 4G, 5E–F; Krug et al. 2013: 1109-1113, figs. 2C, 4; Redfern 2013: 285, fig. 789; Christa et al. 2014: figs. 1C, 3; Krug et al. 2015: 990 -991, figs. 3B, 4.
Type material. Elysia pratensis— Holotype at MCNT (no registration number provided).
Material examined. Bahamas: San Salvador, July 2010, 2 specimens ( LACM 178654–55 View Materials ), Stocking Island , 20 February 2009, 2 specimens ( CPIC 00068–69 ) .
Additional material examined. Florida , USA: Anne’s Beach, Lower Matecumbe Key , June 2007, 1 specimen (isolate Eprat_07Ann01), Geiger Beach, 25 October 2009, 2 specimens (isolate Eprat_09Gei01, isolate Eprat_09Gei01); Discovery Bay , Jamaica, 0 7 March 2006, 1 specimen (isolate Eprat_06Jam01;
Bahamas: Sweetings Cay, July 2007, 26 specimens (isolate Eprat_07Swe01-26), Stirrup Cay, July 2007, 14 specimens (isolate Eprat_07Stir01-14), Little San Salvador, July 2007, 4 specimens (isolate Eprat_07LSS01-04), San Salvador, July 2010, 13 specimens (isolate Eprat_10SSal02, Eprat_10SSal04-15), Bimini, July 2010, 10 specimens (isolate Eprat_10Bim01-10), Compass Cay, July 2010, 4 specimens (isolate Eprat_10Comp01-04), Plana Cays, July 2004, 8 specimens (isolate Eprat_04Pla01-08).
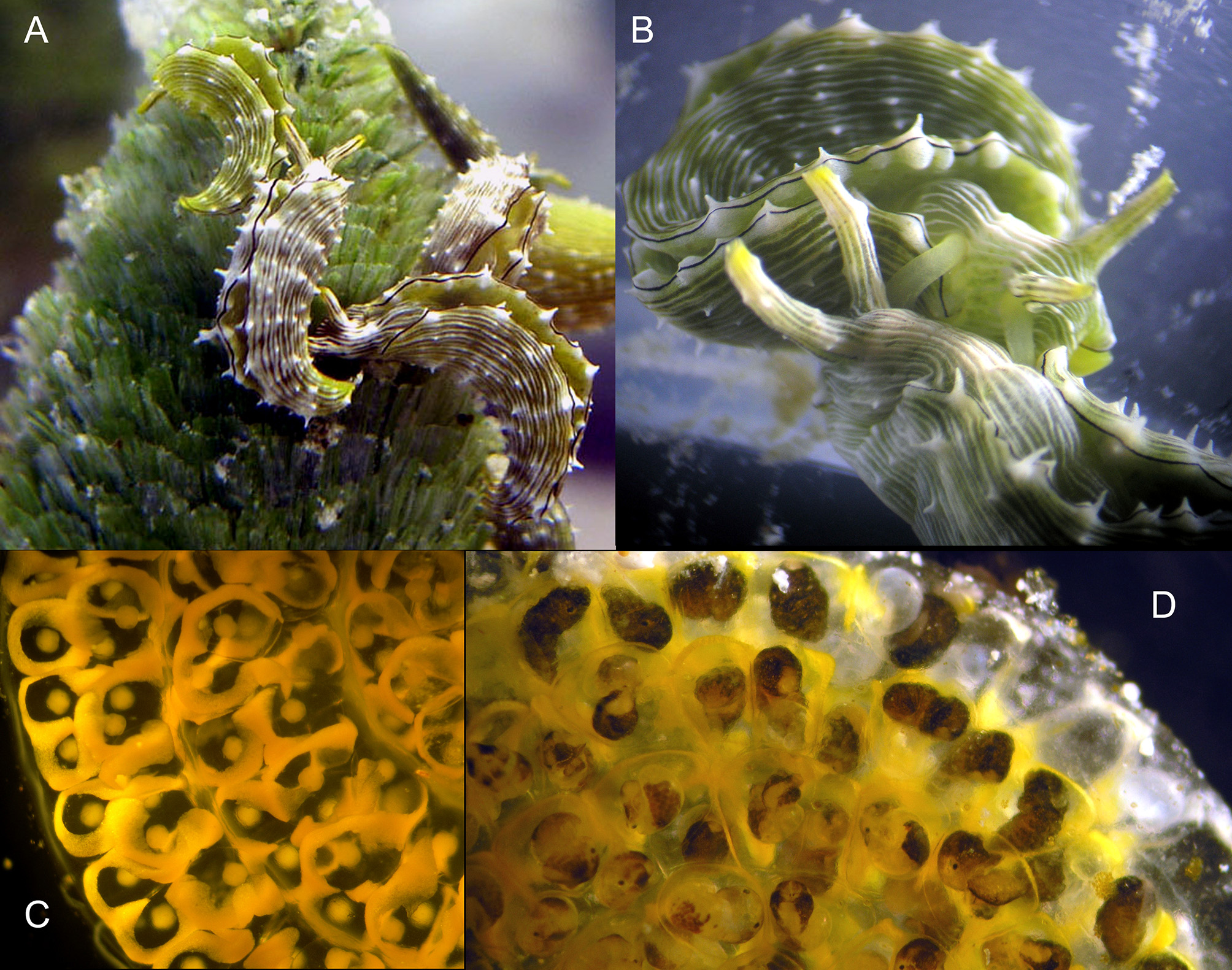
Live animal. Specimens moderately cryptic on host alga, with white longitudinal lines mimicking white borders of individual blades of Rhipocephalus on which slugs have fed, removing the green chloroplasts. Large slugs can be seen atop stipes of the alga in situ, however, due to their large body size and inability to hide within the algal thallus. Parapodia are usually held together to cover dorsum.
External anatomy. Large specimens to 35 mm in body length. Overall coloration green, with longitudinal white stripes running the length of the body along parapodia, over head, and along rhinophores ( Fig. 48 View FIGURE 48 A–B). Body elongated. Head roughly square, eyes small. Rhinophores mostly smooth with a few papillae near distal end. Tips of rhinophores yellow-orange.
Parapodia with black marginal line; on either side, white line with papillae running along edge. Rows of elongated white papillae spaced at regular intervals, emerging from white longitudinal stripes, running length of parapodia. No stripes inside parapodia.
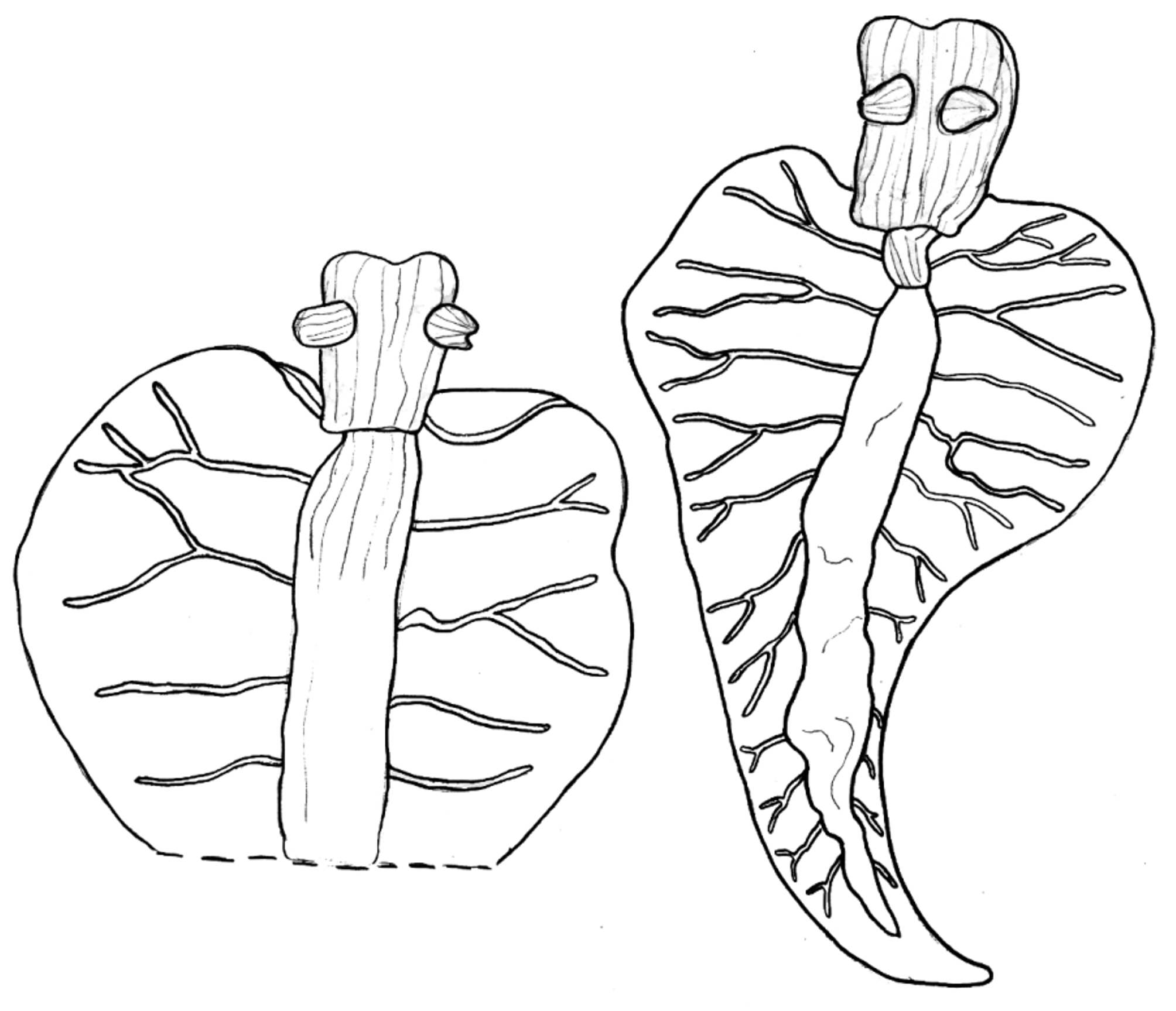
Pericardium green, covered with white longitudinal stripes continuing from head. Renopericardial extension runs entire length of body, with eight or more lateral vessels emerging from along the entire length ( Fig. 49 View FIGURE 49 ). Vessels branch repeatedly and anastomose along inner parapodial margin.
Internal anatomy. Radula with 21 teeth (CPIC 00068), 8 teeth in ascending limb and 13 in descending limb ( Fig. 50 View FIGURE 50 A). Leading tooth elongate and robust (CPIC 00068–69), with 16–22 small, irregular, and coarse denticles on cusp ( Fig. 50 View FIGURE 50 B). Housing depression for interlocking teeth “V”-shaped and extending 80% of tooth length ( Fig. 50 View FIGURE 50 B). Base of tooth approximately ¼ of total tooth length. Ascus containing jumbled heap of discarded teeth ( Fig. 50 View FIGURE 50 C).
Penis elongate with rigid musculature resistant to desiccation (CPIC 00068–69), tapering to an apex devoid of armature. Deferent duct long and highly convoluted ( Figs. 6 View FIGURE 6 R, 50D).
Reproduction and development. Mating was observed for two Florida specimens ( Fig. 48 View FIGURE 48 B). Insemination was largely reciprocal, and similar in all respects to the behaviors described for E. subornata (see Jensen 1986). The penis of each slug contacted its partner between the right anterior parapodial surface and the pericardium for an extended period. Insemination appeared to be hypodermic, with sperm transferred into the epithelial tissue around the pericardium without a penial stylet or puncturing.
A wide flat ribbon of orange ECY weaves around each individual egg capsule ( Fig. 48 View FIGURE 48 C–D). Grand mean clutch size was 146.6 eggs (± 106.4 SE; n = 11 clutches; range = 13 to 1,209 ova per clutch), but only 40.4 eggs ± 6.2 SE if the largest egg mass was excluded. Mean egg diameter ranged from 109.8 µm (± 0.6 SE) to 122.4 µm (± 0.7 SE) for five clutches, with a grand mean egg diameter of 117.9 µm (± 2.2 SE).
Mean larval shell length at metamorphosis was determined for 12 clutches (two from Florida Keys, and five each from Sweetings Cay and Little San Salvador, Bahamas), and ranged from 301.2 µm (± 18.3 SD, n = 20) to 334.5 µm (± 18.2, n = 30); both size extremes were sampled from Little San Salvador parents. Overall, the grand mean shell length was 321.2 µm (± 9.9, n = 12 clutches), close to the median value of 323.4 µm; this is the second largest larval size recorded for a Caribbean sacoglossan, smaller only than E. papillosa . All larvae metamorphosed within the egg mass and emerged as crawl-away juveniles.
Larvae began metamorphosing after 18.4 d (± 0.7 SE, n = 13 clutches) at 22°C, but juveniles required an additional 4.8 d (± 0.7 SE, n = 6) to begin hatching, and about another 5 d for the last juvenile slug to leave the egg mass. Overall, mean developmental time until juveniles began hatching was 23.3 d (± 1.0 SE, n = 6), the longest encapsulated period for any Caribbean elysiid. During the extended post-metamorphic period, juveniles that remained in the egg mass fed readily on ECY, and grew to more than twice the size of siblings that emerged from the egg mass without consuming ECY post-metamorphosis (Krug 2009). Juveniles of E. pratensis were the largest measured for any Caribbean sacoglossan, with a grand mean juvenile length of 651.9 µm (± 42.0 SE, n = 5 clutches).
Host ecology. The host of E. pratensis has not been previously reported; the species takes its name from the seagrass “meadows” from which it was initially discovered by seine netting. Throughout the Caribbean, we have found E. pratensis associated exclusively with the alga Rhipocephalus brevicaulis . Notably, only one small clump of five stipes of R. brevicaulis was found in Discovery Bay, Jamaica, with one large specimen of E. pratensis feeding on the alga. We have observed feeding on R. brevicaulis in the lab; slugs drain cytoplasm from the tips of the filaments which, pressed together, form the scale-like blades. The longitudinal white stripes on E. pratensis render it cryptic against its host, especially when filament tips are white after being fed upon ( Fig. 48 View FIGURE 48 A). Ortea & Espinosa (1996) erroneously presumed the species fed on Thalassia and compared it to other seagrass-feeding species, even discussing the coloration as cryptic on Thalassia ; large, mobile specimens have occasionally been found crawling on other substrates in the field, but feed exclusively upon R. brevicaulis .
Phylogenetic relationships. Elysia pratensis was sister to E. subornata in phylogenetic analyses ( Fig. 4 View FIGURE 4 ), although molecular data were not available for at least one related species ( Elysia hamanni n. sp.); thus, the inclusion of other taxa in subsequent analyses may alter our understanding of relationships in this group. However, a phylogeographic study uncovered hybrid introgression of mitochondria from E. subornata into E. pratensis , resulting in local fixation of mitochondria from E. subornata in northern and central Bahamas populations of E. pratensis ( Rodriguez 2009) . Such cyto-nuclear discordance can lead to misidentification in DNA barcoding studies that employ only mitochondrial markers.
Notably, E. pratensis occupies a highly derived position within subclade 4, in which all other species feed on Caulerpa spp. (Krug et al. 2013). Several features are shared by E. pratensis and other members of the E. tomentosa clade, including a long renopericardium, complex anastomosing network of dorsal vessels, and meandering tubes of bright orange ECY. The derived diet of E. pratensis and its sympatric distribution with E. subornata are consistent with ecological speciation via host shift onto Rhiphocephalus by the ancestor of E. pratensis.
Range. Bahamas (Redfern 2013; present study), Belize (Valdés et al. 2006), Honduras (Valdés et al. 2006), Cuba (Espinosa et al. 2005), Florida, USA, Jamaica (present study), Mexico (Ortea & Espinosa 1996).
Remarks. The longitudinal white stripes distinguish E. pratensis from E. subornata , and the long renopericardial extension running almost the whole body length distinguishes E. pratensis and E. subornata from related elysiids in which the renopericardium runs only partway down the body. There has been little taxonomic confusion surrounding this species; Clark and coworkers appear to have regarded E. pratensis as a variant of E. subornata but undertook no taxonomic study, partly explaining why such a common species was undescribed until relatively recently.
Both diet and pointed, coarsely serrated teeth distinguish E. pratensis from all related and co-occurring species: E. subornata , Elysia pawliki n. sp., and Elysia zemi n. sp. feed on Caulerpa , and have blunt-tipped, nearly smooth teeth. The radula of E. pratensis is presumably adapted to piercing the calcified filaments of R. brevicaulis ; indeed, tooth morphology is strikingly convergent between E. pratensis and the distantly related E. papillosa and E. taino n. sp., which feed on the related and morphologically similar alga Penicillus . In other Caribbean members of the E. tomentosa clade, the descending limb of the radula also forms a pronounced spiral absent from E. pratensis .



| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Elysia pratensis Ortea & Espinosa, 1996
| Krug, Patrick J., Vendetti, Jann E. & Valdés, Ángel 2016 |
Elysia pratensis
| Krug 2015: 990 |
| Redfern 2013: 285 |
| Valdes 2006: 70 |
| Redfern 2001: 163 |
| Ortea 1996: 116 |