
Elysia taino, Krug, Patrick J., Vendetti, Jann E. & Valdés, Ángel, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:91353147-FDA8-45CC-A8F1-1DE801C835A6 |
|
DOI |
https://doi.org/10.5281/zenodo.5664233 |
|
persistent identifier |
https://treatment.plazi.org/id/A04A7E6D-9C3D-FFF4-46C9-F9BAFD2118EC |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia taino |
| status |
sp. nov. |
Elysia taino View in CoL new species
( Figs. 56 View FIGURE 56 G, 70–72)
Elysia papillosa [non Verrill 1901] — Krug et al. 2015: 990-991, figs. 3B, 4
Type material. Discovery Bay, Jamaica, March 2006, (Holotype LACM 3312 View Materials , Paratype LACM 3313 View Materials ) collected by PJK; Stirrup Cay , Bahamas, July 2007 (Paratype LACM 3314 View Materials ), collected by PJK.
Type locality. Discovery Bay, Jamaica.
Material examined. Discovery Bay, Jamaica, March 2006, 5 specimens (Holotype LACM 3312 View Materials , Paratype LACM 3313 View Materials , LACM 178605–07 View Materials ) ; Stirrup Cay , Bahamas, July 2007, 2 specimens (Paratype LACM 3314 View Materials , LACM 178609 View Materials ) .
Additional material examined. Discovery Bay, Jamaica, March 2006, 8 specimens (isolate Etai_06Jam01, isolate Etai_06Jam05-08, isolate Etai_06Jam11-12, isolate Etai_06Jam14) ; Bahamas: Stirrup Cay, July 2007, 3 specimens (isolate Etai_07Stir02, Etai_07Stir08, Etai_07Stir10), Little San Salvador , July 2007, 3 specimens (isolate Etai_07LSS09, Etai_07LSS17, Etai_07LSS19), San Salvador , July 2010, 6 specimens (isolate Etai_10Ssal01, isolate Etai_10Ssal03-07), Plana Cays , July 2007, 8 specimens (isolate Etai_07Pla01, isolate Etai_07Pla04-07, isolate Etai_07Pla10-12), Compass Cay , July 2010, 4 specimens (isolate Etai_10Comp01-04), Northern Exumas , July 2010, 2 specimens (isolate Etai_10NEx01-02), New Providence , July 2010, 5 specimens (isolate Etai_10NPr01-05), Bimini , July 2010, 1 specimen (isolate Etai_10Bim01); Dominica, 2007, 6 specimens (isolate Etai_07Dom01-06) ; US Virgin Islands , 2014, 2 specimens (isolate Etai_14USVI01-02).
Live animal. Specimens swim readily when disturbed by undulating their parapodial margin. Slugs perform hypodermic insemination (sometimes mating in groups) using a long, highly flexible penis that can extend out for half the body length, ~ 10 mm on a large adult specimen.
External anatomy. Coloration and gross external morphology highly variable. Overall color ranging from entirely white ( Fig. 70 View FIGURE 70 A) to mostly light green on the dorsum and base of parapodia, grading to white or brown on the upper parapodial face ( Fig. 70 View FIGURE 70 B–D). Some specimens with longitudinal stripes ( Fig. 70 View FIGURE 70 B) of darker color, and many with scattered tiny brown, red or orange dots. Head and body covered in low white conical papillae. Head generally white to light brown, sometimes with green band under large eyes if white pigment does not cover digestive diverticula. Most specimens with single, large white papilla between eyes; head bisected along midline by row of smaller white papillae in many specimens. Rhinophores relatively short, rolled, with white ground color and rounded white papillae giving a knobby appearance. Dark band appearing about ⅓ of the way up rhinophores, from absence of white pigment and/or concentration of brown-orange dots. Scattered red-orange or brown spots dot head and rhinophores.
Parapodia usually held partially open, not fully covering pericardial complex. Edges of parapodia undulating laterally, producing series of loose siphonal openings along the body. Outer parapodial surface bearing rows of white papillae, varying in size. Lower portion of parapodia adjacent to foot variable in color, ranging from light or dark green to tan, brown or white; distal portion of parapodia typically white on smaller specimens, brown on larger specimens ( Fig. 70 View FIGURE 70 A, C–D). Brick-red or orange flecks scattered over outer parapodial surface ( Fig. 70 View FIGURE 70 A–B).
Parapodial margin with scalloped edge, giving a vertically undulating appearance, due to three-pointed extensions regularly spaced along entire length of margin. Rows of red dots forming thin lines or bars crisscrossing parapodial margin at regularly spaced intervals on most specimens ( Fig. 70 View FIGURE 70 A, D). Ground color of inner parapodial surface and dorsum light to olive green, dotted with white papillae. Dorsum usually pigmented by iridescent blue, green or red-orange speckles, giving glittery appearance. Larger dark green, brown or black spots haphazardly scattered across dorsum. Posterior end of body varying among specimens, sometimes tapering to short, triangular tail, but on other specimens body ending bluntly with no extended tail.
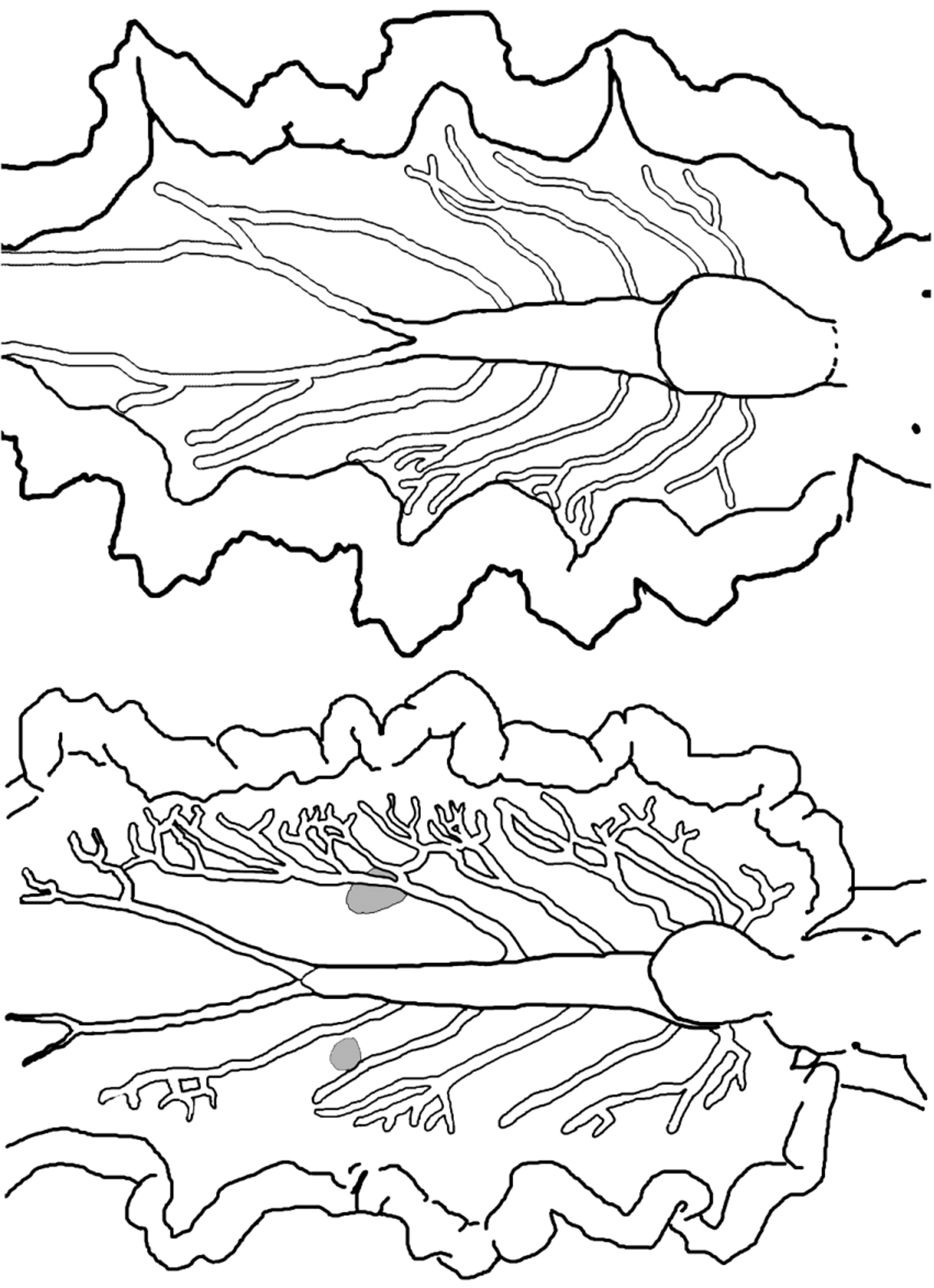
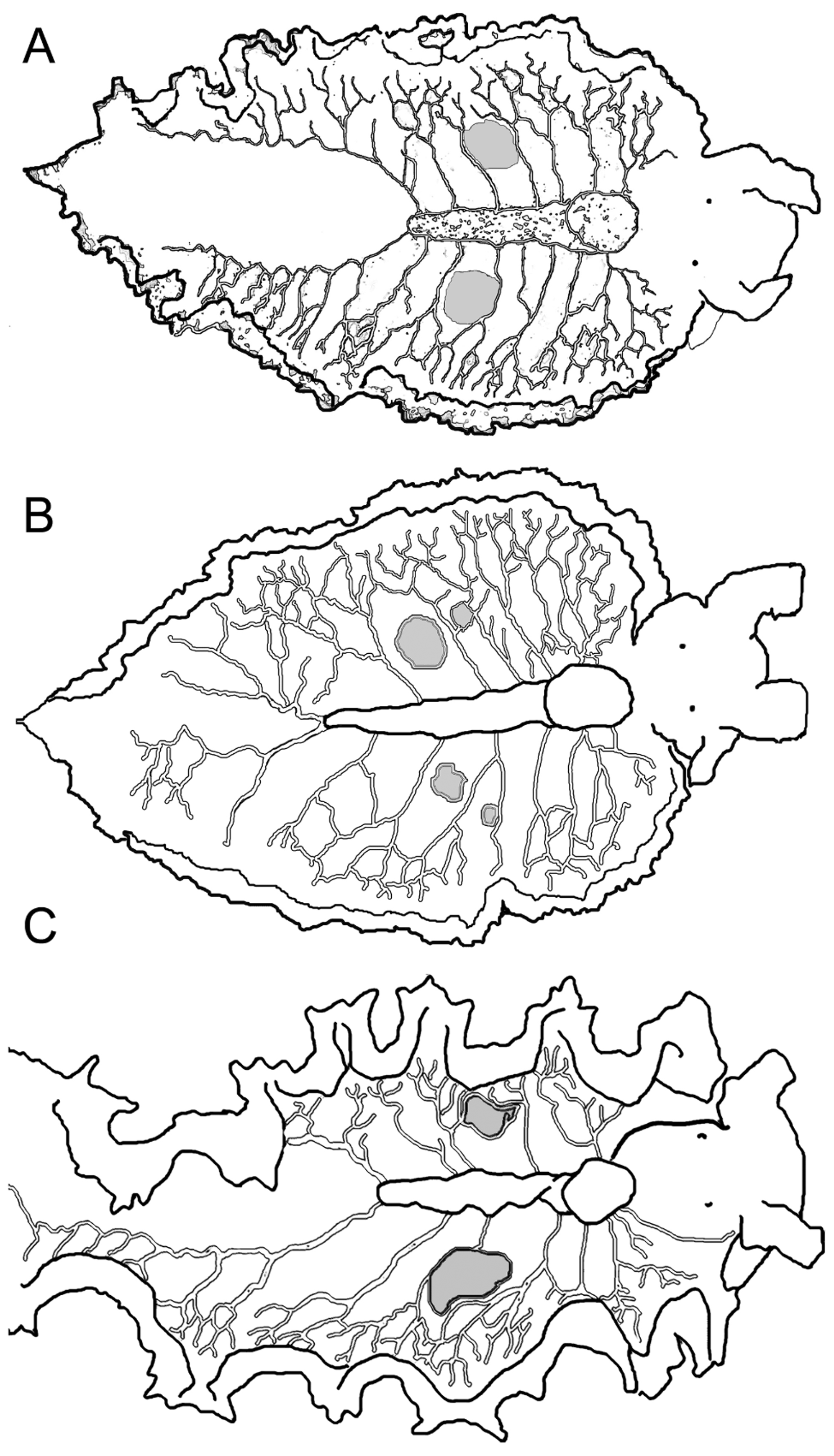
Pericardium small, raised, rounded; white with orange-brown spots ( Fig. 70 View FIGURE 70 C–D). Renopericardial extension distinct from pericardium, clear with orange-brown spots, running less than halfway down dorsal surface; undulating side to side (snake-like) on some larger specimens ( Fig. 70 View FIGURE 70 D). Dorsal vessels often pigmented by iridescent speckles or colored spots creating marked contrast with ground color of dorsal surface (e.g., white vessels against green dorsum, or iridescent blue-green speckles against a white dorsum). Typically 5–6 dorsal vessels emerging on left side and 6–7 on right side of renopericardial complex, with two paired vessels emerging from either side of pericardium and the remaining vessels branching off renopericardial extension ( Figs. 70 View FIGURE 70 C–D, 71). On most specimens, vessels loosely symmetrical in arrangement but with one extra vessel on right side; however vessels highly asymmetrical on other specimens. Vessels wider than in most elysiids, initially straight and unbranched until reaching upper region of inner parapodial surface; vessels then forking or extending thinner side branches at irregular intervals ( Figs. 70 View FIGURE 70 C–D, 71). Terminal branches anastomosing on some specimens but remaining distinct on others. Elongated pair of posterior vessels emerges from terminus of renopericardium, running almost to tail, with numerous anastomosing lateral branches on larger specimens.
One pair of large sperm-storage vesicles typically visible as grey-white swellings of irregular shape ( Figs. 70 View FIGURE 70 D, 71). Sperm vesicles positioned between 3rd and 4th dorsal vessels on either side, about halfway along renopericardial extension, often surrounded on all sides by vessel branches. Vesicles not apparent on juvenile specimens. Upon osmotic shock, swollen vesicles burst releasing sperm.
Size of three specimens: isolate Etain_06Jam17 (LACM 3313), 8.5 mm long × 4.5 mm wide at widest point with parapodia flattened; isolate Etai_06Jam33 (LACM 178605), 7.0 mm long × 4.5 mm wide; isolate Etain_06Jam34 (LACM 178606), 7.5 mm long × 5.0 mm wide.
Internal anatomy. Radula with 11–16 teeth (LACM 3312–3314, LACM 178605–06, LACM 178609), 5–7 in ascending limb and 6–9 in descending limb ( Fig. 72 View FIGURE 72 A, showing only part of ascending limb). Leading tooth elongate and robust bearing 16–24 denticles on cusp ( Fig. 72 View FIGURE 72 B), though one specimen observed with 39 irregular denticles on cusp (LACM 178606). Tooth length, width, and shape somewhat variable. Leading tooth measuring between 120–240 µm long, 20–40 µm wide, with width to length ratio 12.5–20.8 (n=6). Housing depression for interlocking teeth extending ½ the length of tooth ( Fig. 72 View FIGURE 72 B). Base of tooth just less than ½ total tooth length. Ascus containing small, jumbled teeth; not figured (lost during radular preparation).
Penis large and elongate with rigid musculature that did not deform after drying ( Fig. 56 View FIGURE 56 G, 72C), bearing a stylet with a scoop or spoon (LACM 3312–3314, LACM 178605-07) opposite to one medial and two distal flanges ( Fig. 72 View FIGURE 72 D). Deferent duct long, thin, and convoluted.
Reproduction and development. Development was planktotrophic for specimens collected from Jamaica and San Salvador ( Bahamas). Egg masses contain an irregular ribbon of white ECY which contacts most, but not all, egg capsules ( Fig. 70 View FIGURE 70 E). Diameter of uncleaved ova was not determined. Mean larval shell length at hatching was 118.3 µm ± 7.4 (n = 20) for a clutch from Jamaica, and 124.9 µm ± 5.7 (n = 20) for a clutch from San Salvador.
Host ecology. In all field surveys, E. taino n. sp. was found on species of the green algal genus Penicillus (either P. capitatus or P. dumetosus ), on which slugs were also observed to feed in the laboratory. Species-level host preferences are unclear, but in Jamaica, a collection of P. dumetosus (wet weight: 400 g) yielded 34 specimens, while a comparable amount of P. capitatus yielded only 7 specimens. Most specimens on field-collected algae were juveniles <5 mm in body length; in Jamaica, mean length of larger adults was 15.8 mm ± 3.4 SD (n = 9, range = 10–20 mm).
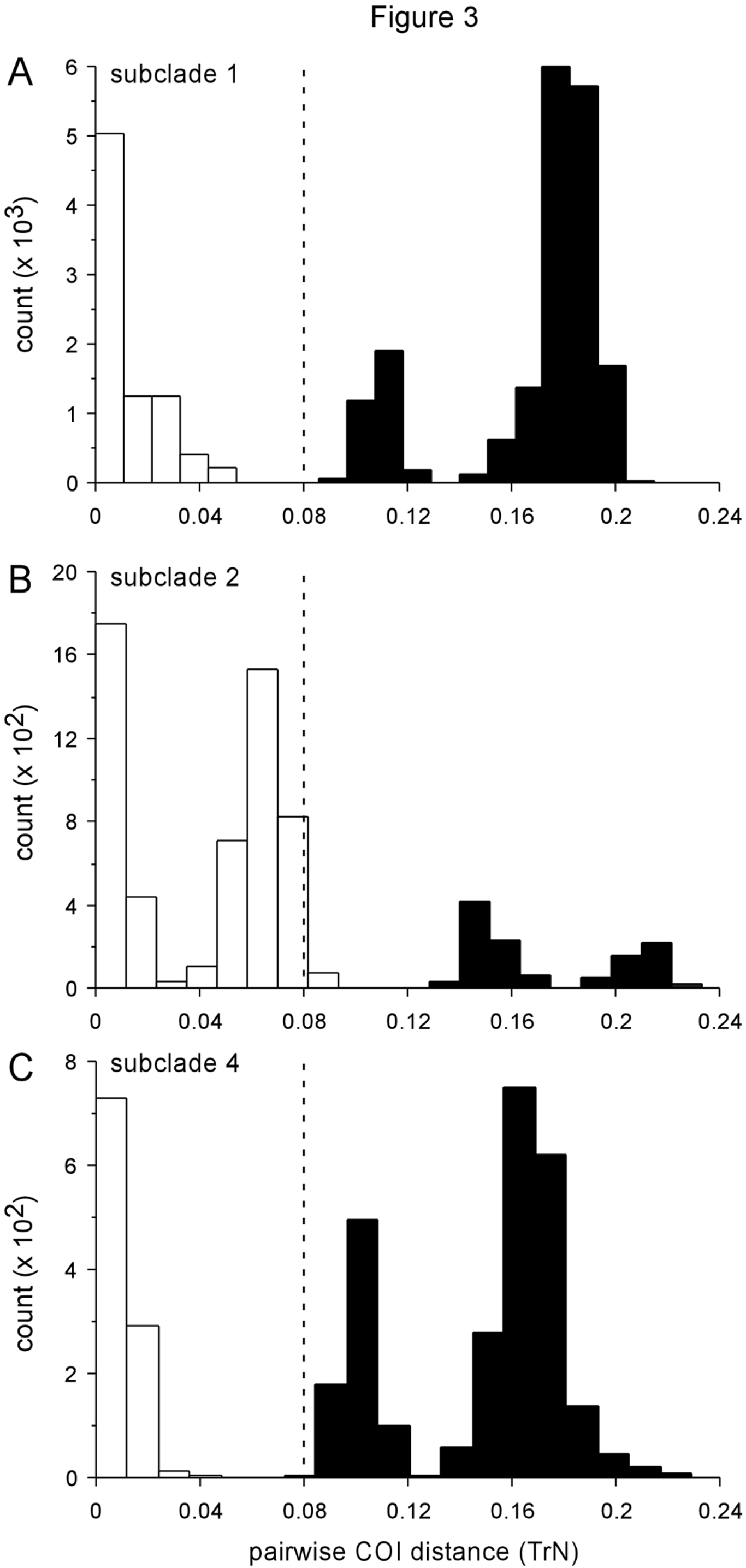
Phylogenetic relationships. Elysia taino n. sp. belongs to subclade 1, and was recovered as sister to the morphologically similar E. papillosa , which also consumes Penicillus ( Fig. 4 View FIGURE 4 ). While host partitioning of lineages onto different algae implicates ecological speciation early in the adaptive radiation of subclade 1, recent divergence between E. taino n. sp. and E. papillosa may have been allopatric as parts of their ranges are non-overlapping. Further study is needed to determine whether E. taino n. sp. and E. papillosa prefer different Penicillus spp. as their host (e.g., P. capitatus versus P. dumetosus ), which could alternatively indicate a role for disruptive selection on host use during the speciation process.
Range. Specimens confirmed through molecular analyses in the present study were from Jamaica, U.S. Virgin Islands, Dominica, and the following Bahamas sites: Sweetings Cay, Stirrup Cay, Little San Salvador, San Salvador, Plana Cays, New Providence, Compass Cay, Northern Exumas, and Bimini.
Etymology. Named in honor of the Taíno , an indigenous people of the Greater Antilles who had a rich mythology of the ocean, and prior to arrival of European colonists were historically distributed throughout the central Caribbean locations where E. taino n. sp. is now common,.
Remarks. The long-unrecognized sister taxon of E. papillosa , E. taino n. sp. was uncovered in our preliminary screen for cryptic species using DNA barcoding. At the COI locus, all sampled E. taino n. sp. specimens (n = 54) formed a monophyletic lineage that was sister to a clade comprising all E. papillosa specimens (n = 120) ( Fig. 3 View FIGURE 3 A; Trathen 2010; authors’ unpublished data). These two clades were a minimum 8.8% divergent.
In comparison, maximum pairwise COI distance within E. taino n. sp. was 3.1%. Moreover, specimens of E. taino n. sp. shared a set of nuclear H3 alleles distinct from the alleles sampled from E. papillosa , even in Jamaica and Bahamas sites where both species co-occur; thus, the two divergent COI lineages are non-interbreeding species, one comprising E. papillosa and the other, E. taino n. sp. Branch lengths from phylogenetic analyses of our fourgene dataset shown in Fig. 4 View FIGURE 4 reflect the species-level divergence between these two taxa.
Elysia taino n. sp. is difficult to distinguish from E. papillosa by morphology, ecology or development. Both species swim when disturbed, have transverse brownish bands on the rhinophores and papillose body surfaces, and possess serrated, straight blade-shaped radular teeth; both feed on Penicillus , have scoop or spoon-like penial stylets, and produce planktotrophic clutches with white ECY. However, molecular data differentiate the two species, and although E. papillosa and E. taino n. sp. co-occur in many localities, parts of their ranges are nonoverlapping. All “ papillosa- like” specimens sampled in Dominica (n=7) and two Bahamas sites (New Providence, Compass Cay) were E. taino n. sp., as were the majority of such specimens sampled in Jamaica (86%) and four Bahamas islands: Stirrup Cay (70%), San Salvador (86%), Plana Cays (80%), and Northern Exumas (67%). In contrast, E. taino n. sp. was the rarer species in three Bahamas sites: Sweetings Cay (6%), Bimini (8%), Little San Salvador (15%). The geographic partitioning of the two species may reflect historical isolation during Pleistocene periods of low sea level, followed by gradual expansion during the Holocene. The lack of admixture at the nuclear H3 locus where the two species are sympatric indicates reproductive isolation has evolved, and thus that the two lineages are biologically good species.
Because no specimens of E. taino n. sp. were collected in Bermuda, this species cannot be the same as E. papillosa Verrill, 1901 . Prior studies on E. papillosa from Curaçao (Ev. Marcus & Er. Marcus 1967) and Florida (Jensen 1980) concerned populations from which E. taino n. sp. has not been sampled, and thus likely comprise valid records of E. papillosa .
The main difference in internal anatomy that permits morphological differentiation is the shape of the radular tooth, which is narrower and more blade-like in E. papillosa versus thicker and more curved in E. taino n. sp. The leading tooth is significantly longer in E. taino n. sp. (188.5 µm ± 21.7 SE; n = 6 specimens) than in comparably sized specimens of E. papillosa (129.7µm ± 11.0 SE; n = 18 specimens) (results of an unpaired two-tailed t test: df = 22, t = 2.58, P = 0.017). Furthermore, the leading tooth is about twice as wide in E. taino n. sp. (30.0 µm ± 3.4 SE; n = 6 specimens) than in E. papillosa (15.6 µm ± 1.6 SE; n = 18 specimens), a highly significant difference (unpaired two-tailed t test: df = 22, t = 4.23, P = 0.0003). Overall, the ratio of tooth width to length is much greater for E. taino n. sp. (16.2 ± 1.1 SE) than in E. papillosa (11.8 ± 0.5 SE), permitting distinction based on radular anatomy that fully accorded with molecular phylogenetic analysis. The angle of curvature for the convex side of the leading tooth is also markedly different between the species. The smooth upper edge of the tooth in E. taino n. sp. angles more sharply from base to tip, making a mean angle of 31.0° ± 3.7 SE. In contrast, the upper surface of the tooth curves only 23.4° ± 1.3 SE in E. papillosa , making the tooth significantly straighter from base to tip (unpaired two-tailed t test: df = 22, t = 2.42, P = 0.025). The tooth of E. papillosa ( Fig. 20 View FIGURE 20 C) also lacks the lateral keel of E. taino n. sp. ( Fig. 72 View FIGURE 72 B).
The dorsal vessel pattern is similar between E. taino n. sp. and E. papillosa , but the vessels may be consistently narrower in E. taino n. sp., and form a more densely anastomosing network along the inner parapodial margin on large specimens ( Fig. 71 View FIGURE 71 ). The sperm-storage vesicles typically form near the middle of the renopericardial extension in E. taino n. sp., but occur near the posterior end of the renopericardial extension in E. papillosa ( Fig. 19 View FIGURE 19 ). Many specimens of E. taino n. sp. also have fine red lines crisscrossing the parapodial margin, a feature not observed in E. papillosa . The same features that differentiate E. papillosa from all other related Caribbean elysiids similarly distinguish E. taino n. sp.





| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.