
Elysia chlorotica Gould, 1870
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:91353147-FDA8-45CC-A8F1-1DE801C835A6 |
|
DOI |
https://doi.org/10.5281/zenodo.5664178 |
|
persistent identifier |
https://treatment.plazi.org/id/A04A7E6D-9C6C-FFA5-46C9-FD83FCAB1A9C |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia chlorotica Gould, 1870 |
| status |
|
Elysia chlorotica Gould, 1870 View in CoL
( Figs. 6 View FIGURE 6 F, 15–17)
Elysia chlorotica Gould 1870: 255 View in CoL –256, pl. 17, figs. 251–255 (Type locality: Cambridge Marshes, Massachusetts) — Verrill 1874: 363, pl. 25, fig. 172; Russell 1946: 96; Pfitzenmeyer 1960: 114; Russell 1964: 37 –38; Bailey & Bleakney 1967: 353; Franz 1970: 7; Ev. Marcus 1972b: 308 –310, fig. 5; Clark 1975: 42; Eyster 1980: 581 –583; Boone 1982: 29 –37, figs. 1–3; Jensen & Clark 1983: 4; Christa et al. 2014, fig. 3; Krug et al. 2015: 990 –991, figs. 3B, 4.
Type material. Elysia chlorotica— unknown location ( Johnson 1964).
Material examined. Martha’s Vineyard, Massachusetts, USA, 2006, 3 specimens ( LACM 178597–99 View Materials ).
Additional material examined. Martha’s Vineyard , Massachusetts, USA, 2006, 8 specimens (isolate Echl_06Mas04-11).
Live animal. Resting slugs often hold parapodia wide open and flattened against the substratum. Slugs have an exceptional ability to osmoconform and hence great tolerance for low salinity, presumably adaptive in their estuarine habitat where salinities can drop rapidly due to runoff from storms ( Pierce et al. 1983, 1984). This is the only species of Elysia that remains green across the entire body surface throughout extended periods of starvation (>1 month), due to prolonged survival of diet-derived chloroplasts.
External anatomy. Overall color deep emerald green; external surfaces dotted with minute speckles of white, light blue and red ( Fig. 15 View FIGURE 15 ). White speckles concentrated into larger white patches spaced at roughly regular intervals across sides of parapodia, and intermediate-sized patches on head ( Fig. 15 View FIGURE 15 A–C). Body surface everywhere smooth with no papillae. Eyespots small, with no other distinguishing markings. Distinct and elongated neck flowing posteriorly into pericardium. Sides of head extend laterally into pointed oral tentacles, whitish at tips ( Fig. 15 View FIGURE 15 B). Rhinophores elongated and tapering to a point, color fading to whitish at tips. Foot not clearly distinct from parapodia, with same overall coloration, narrowing posteriorly.
Wide parapodia covering dorsum when folded up, but often held flat and open by live animal ( Fig. 15 View FIGURE 15 D). Parapodial margin smooth, pale yellow in color from absence of digestive diverticula. Inner parapodial surface emerald green with scattered white and red speckles concentrated along anterior end and margins, and scattered medially along dorsum. Parapodia forming ovoid side flaps, anterior margin with rounded corners; remainder of parapodia equal in width along most of body, gradually tapering to form short triangular tail.
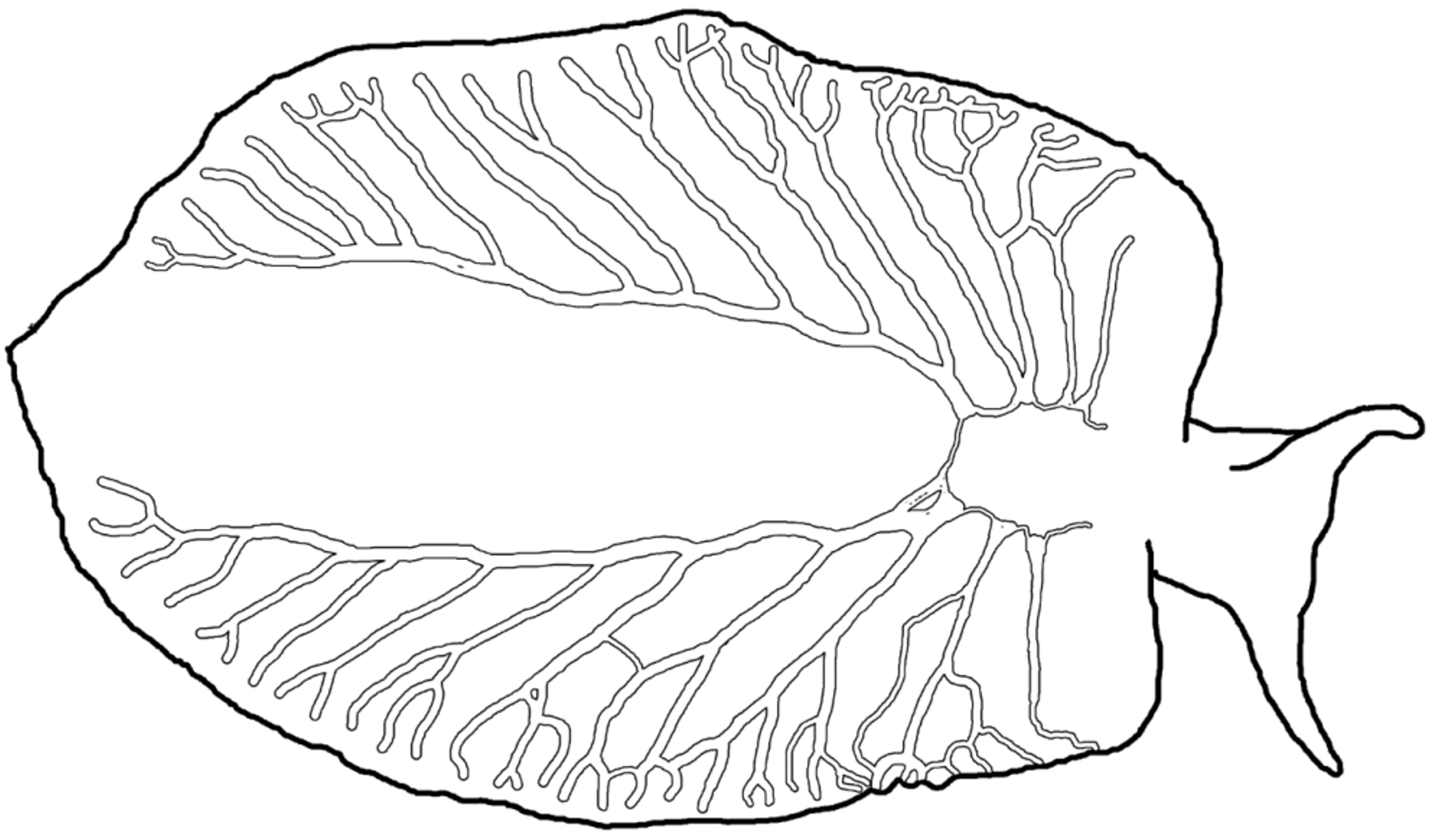
Pericardium round or ovoid, green with dense speckling of white and red dots overlying wider, white renopericardium ( Fig. 15 View FIGURE 15 A–D). Renopericardial extension only slightly posterior of the pericardium, short. Wide, clear dorsal vessels emerging asymmetrically from renopericardial extension, usually three on the right side and three to five on the left side ( Fig. 16 View FIGURE 16 ). Except for posterior pair, vessels fork or send off lateral branches at irregular intervals, side branches sometimes anastomosing; most branches forking immediately before reaching parapodial margin ( Fig. 15 View FIGURE 15 B, D, Fig. 16 View FIGURE 16 ). Posterior vessel on either side immediately forking, then one long branch running whole body length to end of parapodial flaps, and sending off six to nine lateral branches. Vessels transparent, with occasional white or red speckles. Reproductive ducts visible as faint, thin network within dorsal tissue underlying vessel network.
Internal anatomy. Radula with 25 teeth, 13 teeth in ascending limb and 12 in descending limb ( Fig. 17 View FIGURE 17 A). Leading tooth elongate, narrowing to a point, with cusp bearing numerous very small denticles ( Figs. 17 View FIGURE 17 B-C).
Housing depression for interlocking teeth extending approximately 4/5 total tooth length ( Fig. 17 View FIGURE 17 B). Base of tooth approximately ¼ total tooth length. Ascus containing jumbled heap of discarded teeth (not figured).
Penis robust, elongate, and nearly conical with attachment to the body wall, devoid of armature. Deferent duct long, thin, and convoluted ( Fig. 6 View FIGURE 6 F).
Reproduction and development. Like other members of subclade 2, E. chlorotica does not produce ECY. Development is poecilogonous, with alternative larval modes fixed in different populations in Massachusetts, U.S.; all data are taken from West et al. (1984) and errors are SD. Planktotrophic larvae were produced by slugs from Martha’s Vineyard, whereas lecithotrophic larvae with encapsulated metamorphosis were produced by a mainland population at Ipswitch. Planktotrophic clutches contained ~8900 eggs, with a mean diameter of 79 ± 3 µm SD; larvae hatched after 6–7 d at an unspecified temperature, with a mean shell length of 146 ± 11 µm. Larvae cultured on phytoplankton metamorphosed after 2 weeks when presented with either Vaucheria compacta or V. litorea (~50%), suitable adult hosts, with little spontaneous metamorphosis (~1%) occurring on non-host algae ( Enteromorpha or Bryopsis ).
Lecithotrophic clutches contained ~ 175 eggs on average, with a mean diameter of 96 ± 8 mm. Larvae metamorphosed before hatching after 14 ± 1 d (full salinity, 33‰) or 9 ± 1 d (17‰ salinity); faster development at low salinity suggests adaptation to the euryhaline conditions typical of estuaries inhabited by E. chlorotica . Lecithotrophic larval shells were 217 ± 8 µm. Inter-population crosses were successful through two generations, indicating conspecificity. Egg masses of F1 offspring had characteristics similar to their maternal population, while F2 hybrids produced offspring with intermediate characteristics. Despite the rarity of poecilogony, no follow-up studies were published on the lecithotrophic population of E. chlorotica .
Host ecology. Elysia chlorotica feeds on at least two species of the heterokont alga Vaucheria (V. c o m pa c t a and V. litorea ), and may be cultured in the laboratory on the latter ( West et al. 1984; Pelletreau et al. 2012). Although also reported to feed on Cladophora ( Clark 1975) , no supporting data were published. At least three independent radiations onto Vaucheria have thus occurred in Sacoglossa : the genus Alderia , one lineage of Costasiella (2 spp.; Jensen et al. 2014), and E. chlorotica . Most Vaucheria spp. grow in the intertidal zone of estuaries, a stressful environment with low salinities that may impede adaptive shifts onto Vaucheria .
Phylogenetic relationships. Known from the temperate to subtropical northwestern Atlantic, E. chlorotica grouped within subclade 2 as sister to the seagrass-feeding E. serca with strong support. However, the North Atlantic seagrass-feeding E. catulus was not available for phylogenetic analysis, and may be more closely related to E. serca than E. chlorotica , requiring future study.
Range. Nova Scotia ( Bailey & Bleakney 1967; Raymond & Bleakney 1987), Massachusetts ( Gould 1870; Verrill 1874; Russell 1946, 1964), New Jersey ( Franz 1970), Connecticut ( Clark 1975), Maryland ( Pfitzenmeyer 1960), North Carolina ( Abbott 1974), South Carolina ( Eyster 1980), Florida ( Jensen & Clark 1983), Texas ( Boone 1982).
Remarks. Sacoglossans in the clade Plakobranchoidea have long been studied for kleptoplasty, the maintenance of diet-derived plastids in their body tissues for several weeks. A few species exhibit “long-term retention” of plastids for over a month ( Elysia crispata , E. timida , Plakobranchus spp.; Händeler et al. 2009). Exceptionally, E. chlorotica may retain functional chloroplasts for up to nine months, and is the only photosynthetic species with an overall green color due to ramifying digestive diverticula and associated cells harboring plastids (Pierce & Curtis 2012 and references therein). Other long-term retention species may shelter plastids from high sunlight, covering the digestive gland with white or colored parapodia to prevent burn-out of nuclear-encoded proteins from the light-harvesting complex; alternatively, long-term retainers may specialize on algae that still have key genes encoded by the plastid genome (deVries et al. 2013). Regardless, the ability of E. chlorotica to remain green over its entire surface for most of its life is unique even among sacoglossans, prompting special focus on this taxon.
Recent transcriptomic analysis suggested at least 52 genes from the Vaucheria nuclear genome were horizontally transferred into the genome of E. chlorotica , and expressed in adult cells harboring plastids ( Pierce et al. 2009, 2012; Schwartz et al. 2014). Another study failed to detect algal genes in genomic sequences derived from slug eggs, but confirmed that Vaucheria genes were expressed in adult slugs, suggesting algal genes persist as non-integrated or extra-chromosomal DNA after initial feeding ( Bhattacharya et al. 2013). Current evidence is thus consistent with facilitation of long-term retention by expression of algal nuclear genes in E. chlorotica , but the mechanism remains unclear. Recent work showed that after juvenile E. chlorotica were allowed to establish plastid symbiosis for 4 weeks, those held under partial or constant light survived and delayed the loss of body mass for two months without subsequent feeding, whereas slugs held in the dark shrank linearly for a month and then began dying (Pelletreau et al. 2014). Transfer of lipids biosynthesized by photosynthetically active plastids to the slug was key to the establishment and maintenance of kleptoplasty, and to long-term slug survival without food, highlighting a mechanism by which this symbiosis may function.



| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Elysia chlorotica Gould, 1870
| Krug, Patrick J., Vendetti, Jann E. & Valdés, Ángel 2016 |
Elysia chlorotica
| Krug 2015: 990 |
| Jensen 1983: 4 |
| Boone 1982: 29 |
| Eyster 1980: 581 |
| Clark 1975: 42 |
| Ev 1972: 308 |
| Franz 1970: 7 |
| Bailey 1967: 353 |
| Russell 1964: 37 |
| Pfitzenmeyer 1960: 114 |
| Russell 1946: 96 |
| Verrill 1874: 363 |
| Gould 1870: 255 |