
Elysia subornata Verrill, 1901
|
publication ID |
https://doi.org/10.11646/zootaxa.4148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:91353147-FDA8-45CC-A8F1-1DE801C835A6 |
|
DOI |
https://doi.org/10.5281/zenodo.5664185 |
|
persistent identifier |
https://treatment.plazi.org/id/A04A7E6D-9C7F-FFB7-46C9-FB05FD0C1873 |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia subornata Verrill, 1901 |
| status |
|
Elysia subornata Verrill, 1901 View in CoL
( Figs. 6 View FIGURE 6 J, 24–26)
Elysia subornata Verrill 1901: 29 View in CoL –30, pl. 4, fig. 4 ( Type Locality: Castle Harbor, Bermuda) — Pruvot-Fol 1946: 33; Er. Marcus 1957: 414; Ev. Marcus 1980: 66; Clark 1984: 88 –89, figs. 10–14; Hess et al. 1994: 163; Clark 1994: 905; Redfern 2001: 162, figs. 673A–B; Valdés 2006: 66 –67; Krug 2009: 362–365, figs. 4, 6; Händeler et al. 2009: figs. 6–7; Redfern 2013: 285, figs. 790A–C; Zamora-Silva & Ortigosa 2012: 366; Krug et al. 2013: 1109–1113, figs. 2C, 4; Ortigosa et al. 2013: 65; Christa et al. 2014: fig. 3; Krug et al. 2015: 990 –991, figs. 3B, 4.
Elysia cause Er. Marcus 1957: 405 –410, figs. 35–44 ( Type Locality: São Sebastião Island and near Ubatuba, São Paulo, Brazil) — Ev. Marcus & Er. Marcus 1960: 153, fig. 34; Ev. Marcus & Er. Marcus 1963: 22 –23; Er. Marcus & Ev. Marcus 1970: 44 –45, fig. 81; Ev. Marcus & Hughes 1974: 505 –507, figs. 13–14; Thompson 1977: 124, figs. 25h–j, 26f; Clark, Busacca & Stirts 1979: 14 –19, figs.1–7; Jensen & Clark 1983: 4.
Type material. Elysia subornata— untraceable, not at YPMNH; Elysia cauze— possible type specimen, ex. Marcus collection (HMCZ 288301).
Material examined. Anse du Bourg, Terre-de-Haut island , Guadeloupe, January 1986, 1 specimen ( LACM 178629 About LACM ) ; Carriacou island , St. Vincent and the Grenadines (present study), Panama (present study). July 1987, 1 specimen ( LACM 178628 About LACM ) ; Bahamas: Great Exuma, 29 Jan 2009, 1 specimen ( LACM 172297 About LACM ), Stocking Island , 29 Jan 2009, 1 specimen ( CPIC 00076 ), 16 Feb 2009, 1 specimen ( CPIC 00077 ) ; Martinique, June 1986, 1 specimen ( LACM 178631 About LACM ) , July 1987, 1 specimen (LACM 178627), October 2013, 1 specimen (LACM 178630); Water Bay, St. Thomas , U.S. Virgin Islands, 11 April 2006, 1 specimen ( CPIC 00142 ) .
Live animal. Usually found in association with Caulerpa spp. Animal does not swim when disturbed. Parapodia may be held partly open when resting.
External anatomy. Base color ranging from yellow to olive green to dark green. Sides of parapodia pigmented by white to varying degrees; white pigment often arranged in star-shaped clusters around base of white papillae. Scattered tiny black or brown dots all over head and body. Rows of white conical papillae scattered across parapodia and head to varying degrees ( Fig. 24 View FIGURE 24 A–E); some specimens with few papillae ( Fig. 24 View FIGURE 24 B), others densely covered in elongated papillae and associated patches of white pigment ( Fig. 24 View FIGURE 24 D). White, grey or tan patch on top of head starting between rhinophores, bounded by eyes laterally and pericardium posteriorly. White streaks typically extending posteriorly from base of rhinophores over eyes. Anterior end of head trimmed with tan band and dark brown line along the edge. Rhinophores short relative to body length; color tan to lavender to dark brown with rows of white papillae, and white patches concentrated at tips. Parapodia high, covering most of renopericardial complex unless held partly open. Parapodial margin tan to dark brown, sometimes with white speckling. Distinctive fine, black marginal line running along edge of parapodia ( Fig. 24 View FIGURE 24 A–F). Inner surface of parapodia and dorsum green with white speckling ( Fig. 24 View FIGURE 24 B–C, E–F). Posterior end of body narrowing to short, triangular tail, not elongated.
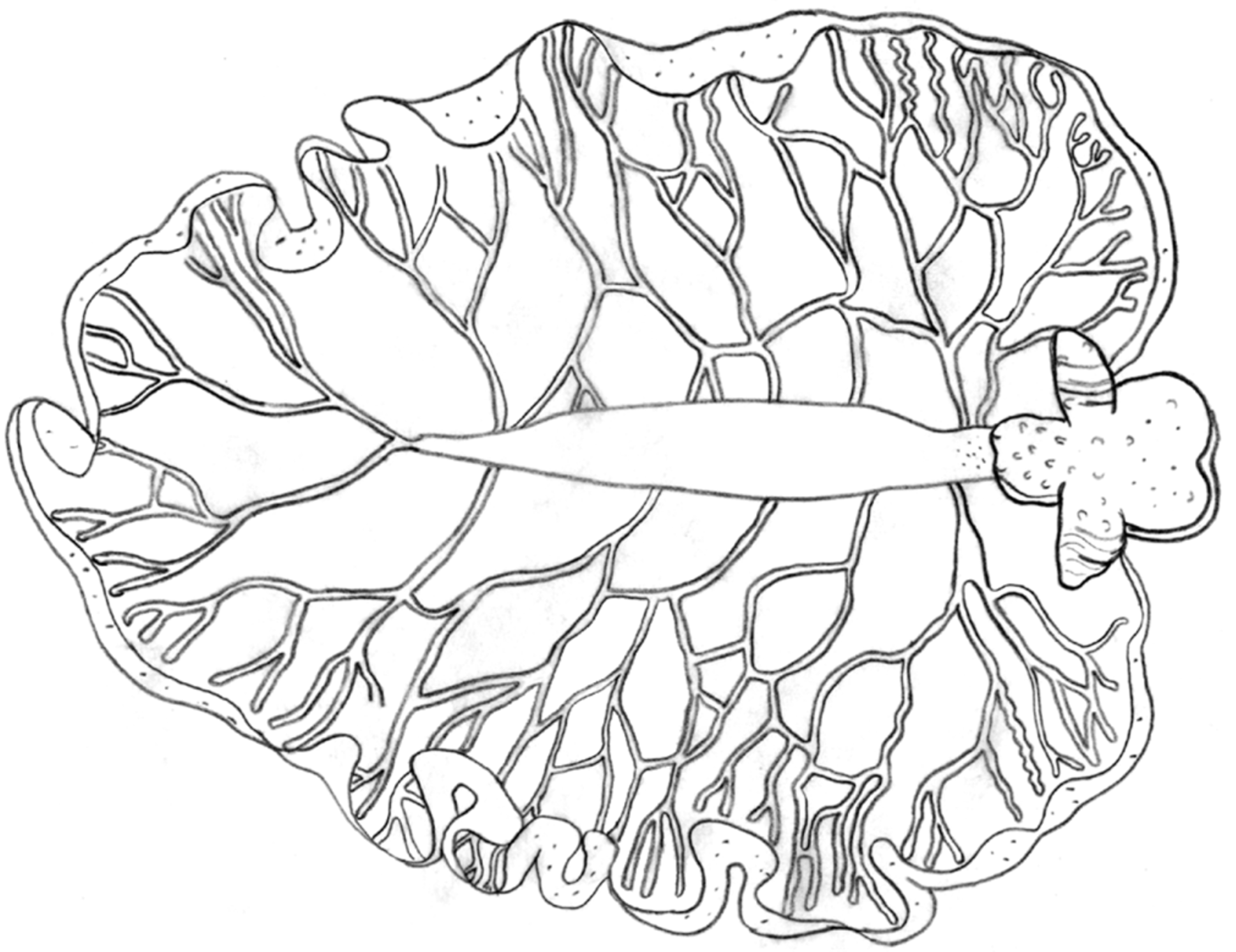
Pericardium small, round, white to brown. Renopericardium light green or whitish, a straight tube running almost entire body length ( Fig. 24 View FIGURE 24 B–C, F). Dorsal vessels clear to whitish, 10 or more emerging on either side of the renopericardium in large specimens ( Figs. 24 View FIGURE 24 F, 25). Vessels relatively symmetric in placement on either side of renopericardium, but branch irregularly and repeatedly; side branches anastomosing into dense network lining upper half of inner parapodial surface ( Fig. 25 View FIGURE 25 ). Posterior vessels not longer than any other vessel pair due to length of renopericardial extension.
Internal anatomy. Radula with 28–29 teeth (LACM 172297, CPIC 0 0 0 76, CPIC 00142), 6 teeth in ascending limb and 21–22 in descending limb ( Fig. 26 View FIGURE 26 A, E). Leading tooth elongate with cusp bearing very fine, blunt denticles. Housing depression for interlocking teeth “V”-shaped and extending ⅔ total tooth length ( Fig. 26 View FIGURE 26 B, F). Base of the tooth approximately ⅓ total tooth length. Ascus with 10 teeth arranged in a short row with some disorganized teeth at the end ( Fig. 26 View FIGURE 26 C).
Penis elongate, often curved (CPIC 00076–77, CPIC 00142) and devoid of armature ( Fig. 6 View FIGURE 6 J, 26D). Deferent duct long, thin, and convoluted.
Reproduction and development. Development is lecithotrophic with 100% encapsulated metamorphosis ( Fig. 24 View FIGURE 24 G–H). Early reports by Clark and colleagues held that E. subornata (as E. cauze ) was poecilogonous, seasonally progressing from planktotrophy (‘ type 1’ development) to swimming lecithotrophic development (‘ type 2’) to encapsulated metamorphosis (‘ type 3’) ( Clark & Goetzfried 1978; Clark et al. 1979). Jensen & Clark (1983) acknowledged that the type 1 and possibly the type 2 egg masses initially attributed to E. subornata were in fact laid by different species, and that E. subornata had encapsulated metamorphosis, the only mode of development we have personally observed.
E. subornata View in CoL produces a continuous string (120–450 µm wide) of bright orange ECY that winds throughout the center of the egg mass, contacting each capsule embedded in the jelly matrix filling the egg spiral ( Fig. 24 View FIGURE 24 G) ( Clark & Goetzfried 1978; Clark et al. 1979; Krug 2009). Embryos develop through a veliger stage with a partially reduced velum, and metamorphose prior to hatching ( Fig. 24 View FIGURE 24 H); a detailed description is provided by Clark et al. (1979). The ECY granules may be absorbed or ingested by larvae during encapsulated development, but can also be consumed directly by juveniles that do not exit the egg mass after metamorphosis; juveniles that remain inside the egg mass and feed on ECY emerge at a significantly larger size than juveniles exiting immediately postmetamorphosis (Krug 2009).
Mean egg diameter for one clutch from the Florida Keys was 119.2 µm (± 2.0 SD; n = 21 ova), matching the 120 µm diameter reported previously ( Clark & Goetzfried 1978; Clark et al. 1979; Clark & Jensen 1981). Mean larval shell length at metamorphosis for two clutches from Florida of 302.0 µm (± 9.5 SD; n = 26) and 311.1 µm (± 12.8 SD; n = 29), again closely matching prior reports of ~300 µm ( Clark et al. 1979). Clark et al. (1979) reported a maximum fecundity of ~1200 eggs per clutch, and time to hatching of 14 d at 23–25°C; Krug (2009) reported a comparable development time of 14.8 d (± 0.9 SD; n = 4 clutches) at 22°C.
Host ecology. Like most members of subclade 4, E. subornata feeds on Caulerpa spp. While other sacoglossans in this group may specialize to some degree on particular species of Caulerpa ( Baumgartner et al. 2009) , E. subornata feeds on at least eight species: C. racemosa ( Jensen 1981b) , C. sertularoides ( Jensen 1981b) , C. paspaloides ( Jensen 1981b) , C. cupressoides ( Jensen 1981b) , C. taxifolia ( Coquillard et al. 2000) , C. mexicana ( Jensen 1981b) and C. ashmeadii ( Clark & Busacca 1978; Clark et al. 1979), and C. verticillata ( Jensen 1981b; Jensen & Clark 1983).
Phylogenetic relationships. In our analyses the sister taxon of E. subornata was E. pratensis ( Fig. 4 View FIGURE 4 ), together forming a clade nested within subclade 4 ( E. tomentosa complex). Species in subclade 4 share an elongated renopericardial complex, which runs almost the full length of the dorsum in both E. subornata and E. pratensis . Molecular data were not available from E. hamanni n. sp., which likely belongs to subclade 4 given its elongated renopericardium and specialization on Caulerpa ; future analyses including this species may alter our understanding of the evolutionary relationships of E. subornata .
Range. Aruba ( Ev. Marcus & Er. Marcus 1963; Valdés et al. 2006), Bahamas ( Redfern 2013), Barbados ( Ev. Marcus & Hughes 1974), Belize ( Clark & DeFreese 1987; Valdés et al. 2006), Bermuda ( Verrill 1901; Clark 1984), Brazil (Er. Marcus 1957), Cayman Islands ( Hess et al. 1994), Bonaire ( Ev. Marcus & Er. Marcus 1963); Curaçao ( Ev. Marcus & Er. Marcus 1963; Er. Marcus & Ev. Marcus 1970), Florida ( Ev. Marcus & Er. Marcus 1960; Er. Marcus & Ev. Marcus 1970; Jensen & Clark 1983; Clark 1994) , Guadeloupe (present study), Jamaica ( Thompson 1977), Martinique ( Thibaut et al. 2001), Mexico ( Valdés et al. 2006; Zamora-Silva & Ortigosa 2012; Ortigosa et al. 2013), Puerto Rico ( Er. Marcus & Ev. Marcus 1970), St. Martin / St. Maarten ( Ev. Marcus & Er. Marcus 1963; Er. Marcus & Ev. Marcus 1970), St. Thomas , U.S. Virgin Islands (present study) , St. Vincent and the Grenadines (present study), Virgin Islands ( Valdés et al. 2006).
Remarks. Clark (1984) synonymized Elysia cauze with Elysia subornata . Clark indicated that different species with planktotrophic and pelagic lecithotrophic development were initially misidentified as E. subornata , leading to the erroneous reports of seasonally varying development mode in this species; however, the identities of those other two species were never published. Clark’s notes indicate he took E. pratensis to be an ecotype of E. subornata , but as E. pratensis also has lecithotrophic development with encapsulated metamorphosis, it cannot be one of the misidentified species with swimming larvae. The elongated renopericardium distinguishes E. subornata from all other Caribbean species except E. pratensis . Radular characters and host use distinguish E. subornata from E. pratensis : E. subornata has nearly smooth teeth and feeds on Caulerpa spp., whereas E. pratensis has coarsely serrated teeth and feeds on Rhipocephalus phoenix .




| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heterobranchia |
|
SuperOrder |
Sacoglossa |
|
Order |
|
|
Family |
|
|
Genus |
Elysia subornata Verrill, 1901
| Krug, Patrick J., Vendetti, Jann E. & Valdés, Ángel 2016 |
Elysia cause
| Jensen 1983: 4 |
| Clark 1979: 14 |
| Thompson 1977: 124 |
| Ev 1974: 505 |
| Er 1970: 44 |
| Ev 1963: 22 |
| Ev 1960: 153 |
Elysia subornata
| Krug 2015: 990 |
| Redfern 2013: 285 |
| Ortigosa 2013: 65 |
| Zamora-Silva 2012: 366 |
| Valdes 2006: 66 |
| Redfern 2001: 162 |
| Hess 1994: 163 |
| Clark 1994: 905 |
| Clark 1984: 88 |
| Ev 1980: 66 |
| Pruvot-Fol 1946: 33 |
| Verrill 1901: 29 |